|
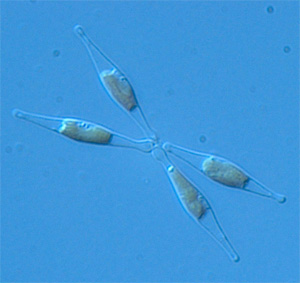
Диатомовые водоросли появились сравнительно недавно по геологическим масштабам времени, однако успели стать одной из важнейших групп фотосинтезирующих организмов на планете. Анализ генома диатомеи Phaeodactylum показал, что удивительный эволюционный успех диатомей во многом объясняется их способностью заимствовать полезные гены у бактерий.
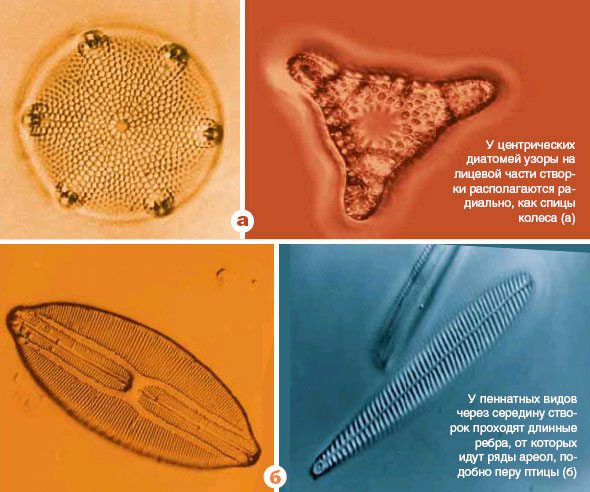
Большая международная группа ученых сообщила в последнем номере журнала Nature о прочтении генома диатомовой водоросли Phaeodactylum tricornutum. Это уже второй вид диатомовых водорослей, геном которого удалось прочесть (первым в 2004 году был опубликован геном диатомеи Thalassiosira). Диатомеи подразделяются на центрических и пеннатных (см. рисунки), причем Thalassiosira относится к первой группе, а Phaeodactylum — ко второй.Интерес исследователей к диатомеям далеко не случаен: по примерным оценкам, на долю этих одноклеточных водорослей приходится около 20% всей первичной продукции биосферы. Диатомеи практически вездесущи: их можно встретить не только во всех морях и пресных водоемах, но и в таких экстремальных местообитаниях, как высокогорные пустыни и антарктические льды.
Диатомеи — группа сравнительно молодая. Древнейшие находки ископаемых центрических диатомей известны из отложений юрского периода (около 180 млн лет назад). Пеннатные диатомеи вдвое моложе: они известны лишь начиная с позднего мела (90 млн лет назад); при этом они гораздо разнообразнее и многочисленнее своих центрических предков.
|
Имея в своем распоряжении два полных генома диатомей, ученые смогли сделать ряд важных выводов об эволюции группы.
Сравнительная характеристика геномов двух видов диатомей:
Пеннатная диатомея Phaeodactylum
Центрическая диатомея Thalassiosira
Размер генома
27,4 млн пар нуклеотидов
32,4 млн пар нуклеотидов
Число генов
10402
11776
Число генов, встречающихся во многих группах эукариот
3523
4332
Число генов, встреченных у обеих диатомей и больше ни у кого (цифры в этой строчке не одинаковы из-за различного числа генных дупликаций у двух видов диатомей)
1328
1407
Число генов, уникальных для данного вида
4366
3912
Число интронов (некодирующих вставок в генах)
8169
17880
Доля мобильных генетических элементов — ретротранспозонов класса LTR
5,8%
1,1%
Один из основных выводов состоит в том, что эволюция диатомей происходила на удивление быстро. Лишь для 57% генов пеннатной диатомеи нашлись гомологи (близкородственные гены) в геноме центрической диатомеи. Аминокислотные последовательности белков у двух диатомей совпадают только на 54,9%. Для сравнения, белки человека и рыбы фугу имеют 61,4% одинаковых аминокислот, человека и асцидии — 52,6%. Таким образом, по строению белков две диатомеи сильнее отличаются друг от друга, чем человек от рыбы, но меньше, чем человек от низших хордовых, представителем которых является асцидия. Между тем эволюционные линии млекопитающих и лучепёрых рыб разошлись, по имеющимся оценкам, около 450 млн лет назад, то есть за много сотен миллионов лет до появления диатомей. Это значит, что темпы молекулярной эволюции у диатомей были в несколько раз выше, чем у позвоночных.
Дупликации (удвоения) больших фрагментов генома не играли в эволюции диатомей такой важной роли, как у позвоночных (см.: Геном ланцетника помог раскрыть секрет эволюционного успеха позвоночных, «Элементы», 23.06.2008). Особенностью эволюции центрических диатомей было приобретение (по-видимому, сравнительно недавнее) большого количества новых интронов — некодирующих вставок в генах. У пеннатных диатомей массового распространения интронов не было, зато в их геномах бурно размножились мобильные генетические элементы — ретротранспозоны (см. таблицу). Проанализировав дополнительные данные по другим видам пеннатных диатомей, авторы пришли к выводу, что активность ретротранспозонов была важным фактором, способствовавшим росту видового разнообразия этой группы. Перепрыгивая с место на место внутри генома, ретротранспозоны могут влиять на активность соседних генов и способствовать росту генетической изменчивости.
Диатомеи вместе с бурыми и золотистыми водорослями и некоторыми другими эукариотами входят в состав группы Heteroconta. Считается, что гетероконты появились около 1 млрд лет назад в результате симбиоза гетеротрофного (нефотосинтезирующего) одноклеточного организма с одноклеточной же красной водорослью. У красных водорослей, как и у зеленых растений, хлоропласты (органеллы, служащие для фотосинтеза) являются первичными, то есть происходят напрямую от симбиотических цианобактерий. Первичные хлоропласты всегда окружены двумя мембранами. Предки гетероконт проглотили одноклеточную красную водоросль и превратили ее в фотосинтезирующего симбионта. Впоследствии от клетки красной водоросли почти ничего не осталось, кроме внешней оболочки и хлоропласта. Гены симбиотической красной водоросли были отчасти утрачены, отчасти — перешли в геном хозяина. Поэтому хлоропласты гетероконт называют «вторичными», и они окружены не двумя, а четырьмя мембранами (из них две внутренние — это оболочка первичного хлоропласта, третья изнутри — бывшая оболочка клетки красной водоросли, а четвертая, внешняя — это оболочка пузырька-вакуоли, в которую была заключена проглоченная красная водоросль).
Авторы проверили эту теорию, предприняв целенаправленный поиск в геномах диатомей генов, похожих на гены красных водорослей. Поиск увенчался успехом: удалось выявить более 170 генов, унаследованных предками диатомей от красных водорослей. Подавляющее большинство этих генов необходимо для работы хлоропластов. Этот результат — очень весомый довод в пользу того, что сложившиеся на сегодняшний день представления о ранней эволюции эукариот в общих чертах верны.
В геномах диатомей нашлось также большое количество уникальных генов, аналогов которых нет у других живых организмов. Многие из этих новых генов возникли в результате дублирования и перетасовки фрагментов старых генов. По-видимому, важной движущей силой этих перекомбинаций была деятельность мобильных генетических элементов — ретротранспозонов.
|
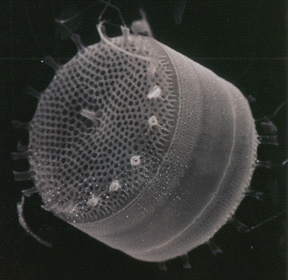
Самое удивительное, что в геномах диатомей обнаружилось очень много генов, которые явно были заимствованы диатомеями у различных прокариот: цианобактерий, протеобактерий, архей и других. В геноме Phaeodactylum обнаружено 587 таких заимствованных генов. На сегодняшний день это рекордное количество генов прокариотического происхождения, найденных в эукариотическом геноме. Более половины из этих генов (56%) есть также и у Thalassiosira. Эти гены, скорее всего, были заимствованы диатомеями у бактерий довольно давно — еще до расхождения эволюционных линий центрических и пеннатных диатомей. Остальные 44%, по всей видимости, были заимствованы предками Phaeodactylum уже после этого события, то есть в течение последних 90 млн лет.
Бактериальные гены, по-видимому, значительно расширили биохимические возможности диатомей. Эти гены помогают диатомеям осуществлять ряд биохимических реакций, не свойственных другим эукариотам. Кроме того, они участвуют в построении ажурных кремневых раковинок — главной «визитной карточки» диатомей, которая во многом обеспечила их эволюционный успех. Диатомеи «одолжили» у бактерий также и многие гены рецепторных и сигнальных белков, при помощи которых бактерии воспринимают сигналы из окружающей среды и реагируют на них. Среди заимствованных у бактерий рецепторов есть даже несколько светочувствительных белков, благодаря которым диатомеи могут реагировать на изменения освещенности.
Авторы предполагают, что активный обмен генами между диатомеями и бактериями был одной из главных причин быстрой эволюции диатомей и их эволюционного успеха. Полученные результаты говорят о том, что горизонтальный генетический обмен, по-видимому, играет в эволюции эукариот (по крайней мере одноклеточных) более важную роль, чем считалось до сих пор.

")