|
Одна из важнейших задач, стоящих перед эволюционной биологией, состоит в расшифровке генетических механизмов, ответственных за формирование сложных признаков. Идеальный объект для таких исследований — плодовые мушки рода Drosophila. Это едва ли не самая хорошо изученная группа животных, включающая множество видов с разнообразной морфологией, причем характер родственных связей между видами в большинстве случаев установлен достаточно надежно (то есть известно, кто от кого и когда произошел).
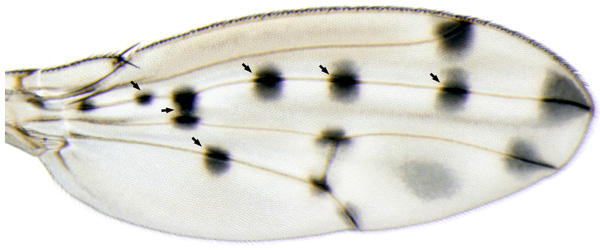
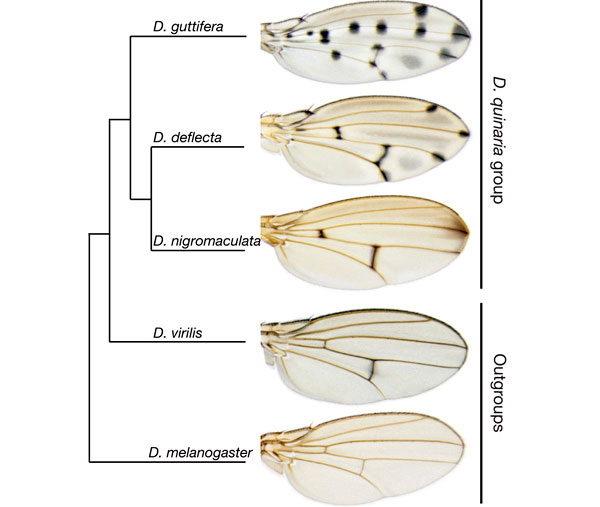
Многие морфологические признаки дрозофил быстро меняются в ходе эволюции. При этом иногда происходит явное усложнение морфологии. Например, у D. guttifera на крыльях имеется замысловатый узор, которого нет у других дрозофил. Узор состоит из 16 черных пятен, расположенных в строго определенных местах на продольных жилках и в точках соединения продольных жилок с поперечными, и нескольких серых «теней» между жилками. У всех ближайших родственников D. guttifera крылья либо вовсе не окрашены, либо имеют гораздо более простой орнамент. По-видимому, сложная окраска крыльев D. guttifera является недавним эволюционным приобретением.
Шон Кэрролл (Sean B. Carroll), известный американский генетик и автор научно-популярных книг, и его коллеги из Висконсинского университета (University of Wisconsin) решили выяснить, какие генетические изменения обеспечили появление у D. guttifera затейливого орнамента на крыльях. Ранее эта исследовательская группа установила, что появление простых темных пятен на крыльях у разных видов дрозофил было связано с модификациями регуляторных участков гена yellow, который управляет синтезом черного пигмента меланина и отвечает за пигментацию разных частей тела. Об этом исследовании рассказано в заметке Тонкая подстройка многофункционального гена может приводить к появлению новых признаков («Элементы», 25.04.2006).
Опираясь на эти результаты, авторы начали свой поиск с изучения работы гена yellow в формирующихся крыльях D. guttifera на поздних стадиях развития куколки. Оказалось, что yellow работает как раз в тех участках крыла, где впоследствии появятся темные пятна. Следовательно, у этого вида, как и у других дрозофил, формирование пятен идет под управлением yellow. Однако у D. guttifera распределение областей экспрессии yellow по крыловой пластинке отличается особой сложностью.
Следующая задача, стоявшая перед авторами, заключалась в расшифровке механизмов, которые заставляют yellow работать в одних частях крыла и не работать в других. Ранее было установлено, что узоры на крыльях дрозофил зависят от некодирующих регуляторных участков гена yellow. Авторы отсеквенировали фрагмент генома D. guttifera длинной в 42 тысячи пар нуклеотидов, включающий ген yellow и его окрестности. Затем они подразделили некодирующие области отсеквенированного участка на 28 перекрывающихся кусочков и стали изучать их свойства при помощи генно-инженерных экспериментов. Каждый кусочек по очереди присоединяли к гену зеленого флуоресцирующего белка (enhanced green fluorescent protein, eGFP) и вставляли эту конструкцию в геном D. guttifera, а затем смотрели, где будет вырабатываться eGFP. Таким непростым способом удалось выявить регуляторный участок длиной в 277 пар нуклеотидов, который заставляет присоединенный к нему ген включаться как раз в тех 16 точках развивающегося крыла, где у диких мух D. guttifera включается ген yellow и формируются пятна на жилках. Этот регуляторный участок находится на расстоянии около 5 тысяч пар нуклеотидов перед началом кодирующей части yellow. Таким образом, один-единственный регуляторный элемент контролирует формирование всех 16 пятен.
|
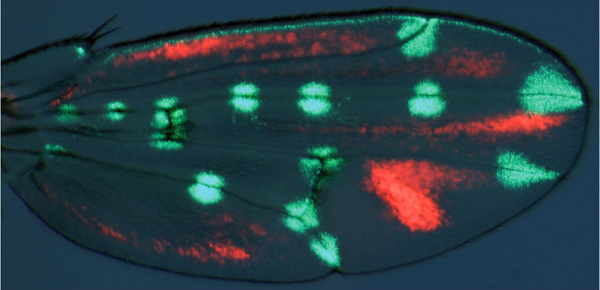
Тем же способом был выявлен еще один регуляторный участок, который отвечает за серые «тени» между жилками. Этот участок длиной в 414 пар нуклеотидов расположен в первом интроне гена yellow. Если присоединить его к гену светящегося белка и вставить в геном D. guttifera, у куколки начинают светиться те участки крыла, где должны быть «тени».
Следовательно, всего лишь два регуляторных элемента определяют всё распределение областей экспрессии yellow по крыловой пластинке (хотя в принципе каждое пятнышко могло бы определяться своим собственным регуляторным элементом). Первый из обнаруженных элементов авторы назвали vein spot (vs), второй — intervein shade (iv). Эти элементы действуют как переключатели. Первый из них срабатывает там, где должны сформироваться пятна, второй — в местах образования будущих «теней».
Таким образом, «нижняя» часть регуляторного каскада была в общих чертах расшифрована. Распределение пятен по крылу определяется элементами vs и iv. Срабатывание переключателей vs и iv активирует ген yellow. Ген yellow включает синтез меланина. Синтез меланина приводит к формированию пятна на крыле.
Этого уже было бы достаточно для хорошей статьи. Но авторы не остановились на достигнутом и продвинулись еще на один шаг «вверх» по регуляторному каскаду. Им удалось выяснить, на какой сигнал реагирует переключатель vs — что заставляет его включаться в нужном месте и в нужное время (систему управления переключателем iv авторы в данной статье не рассматривают — ведь нужно что-то оставить и на потом).
Не имея никаких «подсказок», подступиться к такой задаче непросто, ведь в принципе присоединяться к элементу vs, запуская экспрессию yellow, мог бы любой из многих сотен регуляторных белков (факторов транскрипции), а на работу этого неизвестного белка наверняка влияют еще какие-то факторы. Авторы, однако, сумели получить важную подсказку, изучая разнообразные отклонения от нормального хода развития в многочисленной лабораторной популяции мух D. guttifera. Наблюдения показали, что пятна жестко привязаны к определенным ориентирам или «опорным точкам» (landmarks) на крыловой пластинке. Такими опорными точками являются поперечные жилки и места их соединения с продольными жилками, окончания продольных жилок, а также колоколовидные сенсиллы — механорецепторы, расположенные на продольных жилках. Если у мутантной мухи на крыле появляется новая сенсилла или новое место соединения жилок, это всегда приводит к формированию нового пятна. Если, наоборот, сенсилла теряется, то исчезает и соответствующее пятно.
Следовательно, сложный рисунок на крыльях тесно связан с предсуществующей «разметкой» крыловой пластины, которая определяет ее структуру. Это позволило ограничить круг поиска. Генетическая разметка развивающегося эмбриона или куколки осуществляется сравнительно небольшим набором генов — ключевых регуляторов развития (подробнее см. в «ответе на детский вопрос» о принципах биологии развития и в заметке Эволюция ящериц и змей сопровождалась изменениями Hox-генов, «Элементы», 15.03.2010). Возможно, какой-то из этих морфогенов (см. Morphogen) и взаимодействует — прямо или опосредованно — с переключателем vs.
Чтобы найти искомый ген, авторы пересадили элемент vs, присоединенный к гену eGFP, в геном мухи D. melanogaster — классического лабораторного объекта, чья генетика развития изучена гораздо лучше, чем у D. guttifera. Оказалось, что у D. melanogaster элемент vs срабатывает у основания крыла и на поперечных жилках, но не работает вблизи колоколовидных сенсилл. Точно так же распределены у этого вида и области экспрессии гена wingless — одного из важнейших регуляторов индивидуального развития (см. Wnt signaling pathway). На ранних этапах онтогенеза членистоногих wingless участвует в глобальной разметке туловища, формировании конечностей, определяет границы и полярность сегментов (см.: Конечности у членистоногих и кольчатых червей растут из общего корня, «Элементы», 20.11.2008). На поздних стадиях развития куколки wingless занят менее грандиозными проектами, участвуя в том числе и в оформлении некоторых деталей строения крыльев.
Пересаженный от D. guttifera элемент vs сработал в крыле куколки D. melanogaster ровно в тех местах, где активен ген wingless. Это делает wingless хорошим кандидатом на роль «входного сигнала», вызывающего активизацию vs и включение yellow. Авторы проверили, как распределены области экспрессии wingless в крыле D. guttifera, и убедились, что всё сходится: у вида с пятнистыми крыльями wingless на стадии куколки работает как раз в тех участках крыла, где позже формируются пятна.
|
Чтобы окончательно убедиться в том, что активность wingless является достаточным условием для появления пигментных пятен на крыльях у D. guttifera, авторы провели еще одну серию генно-инженерных экспериментов. На этот раз в эмбрионы D. guttifera впрыскивали ген wingless, взятый у D. melanogaster и соединенный с регуляторным участком, заимствованным у дрожжей. Смысл этой деятельности заключался в том, чтобы получить трансгенных мух D. guttifera, у которых области экспрессии wingless в крыльях были бы расположены как-то иначе. В конце концов такие мухи были получены: у них возникла новая область экспрессии wingless вдоль одной из продольных жилок (L4). В полном соответствии с теоретическими ожиданиями у этих мух появилась и новая темная полоса на крыльях, идущая вдоль жилки L4.
Авторы также проанализировали пигментацию крыльев, экспрессию wingless и наличие или отсутствие регуляторного элемента vs у других видов дрозофил, что позволило восстановить ход эволюции узора на крыльях.
|
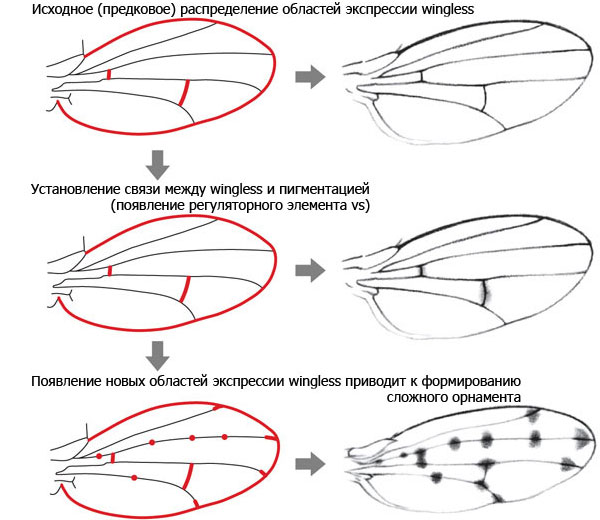
Исходно у дрозофил ген wingless экспрессировался вдоль поперечных жилок, но не работал у концов продольных жилок и в колоколовидных сенсиллах. В гене yellow не было регуляторного элемента vs, способного реагировать на активность гена wingless, а крылья не имели орнамента. Это исходное состояние сохранилось у D. melanogaster. Затем у общего предка групп D. virilis и D. quinaria появился элемент vs, что привело к формированию связи между экспрессией wingless и пятнами на крыльях. Первые темные пятна поэтому появились там, где исходно экспрессировался wingless — вдоль поперечных жилок.
После того, как пигментация стала зависеть от wingless, у эволюции появилась возможность создавать новые узоры, меняя экспрессию этого гена. wingless имеет сложную и пока еще слабо изученную систему регуляции, но в целом его работа ориентируется на базовую морфогенетическую «разметку» и ключевые «опорные точки» развивающегося организма. Эта привязанность гена wingless к «опорным точкам» сохранилась и в ходе дальнейших изменений его экспрессии в крыльях. У некоторых видов группы quinaria появились новые области экспрессии wingless на концах продольных жилок, что сопровождалось формированием соответствующих пятен. Только у одного вида — D. guttifera — орнамент крыльев усложнился еще сильнее за счет появления дополнительных областей экспрессии wingless в шести колоколовидных сенсиллах.
Получается, что замысловатый орнамент крыльев D. guttifera развился на основе существовавшего ранее сложного плана строения крыла. Авторы предполагают, что примерно таким же образом, отталкиваясь от «опорных точек» морфогенетической разметки крыла, сформировались узоры на крыльях и у других насекомых, включая бабочек. Но чтобы это доказать, придется еще немало потрудиться.
Работа представляет собой важный шаг к пониманию генетических механизмов формирования новых сложных признаков. Кроме того, она наглядно показывает, как мало мы еще знаем об этих механизмах и как много нерешенных задач стоит перед молодой научной дисциплиной, призванной их решить — эволюционной биологией развития, или evo-devo.

")