Излюбленным аргументом противников Дарвина с давних пор была кажущаяся неспособность его теории объяснить происхождение камбалообразных рыб. Утверждалось, что перемещение глаза с одной стороны головы на другую не могло происходить постепенно, потому что на начальных этапах эти изменения не приносили бы никакой пользы своим обладателям. Однако найденные в эоценовых отложениях Италии и Франции ископаемые переходные формы в очередной раз подтвердили правоту дарвиновской теории.
|

|
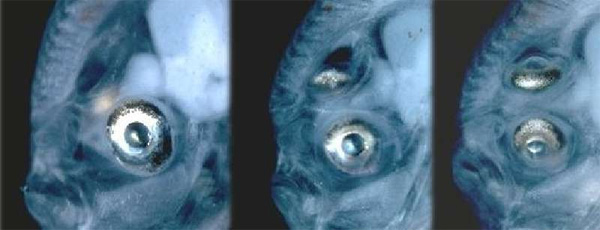
Дарвин и его последователи предполагали, что изменения в строении черепа, наблюдающиеся у камбалообразных в ходе индивидуального развития, воспроизводят эволюцию группы. Не зная материальной природы наследственности и изменчивости, Дарвин допускал, что в данном случае мог работать «ламарковский» эволюционный механизм: предкам камбал, повадившимся лежать на боку, приходилось постоянно косить глаза, и прижизненные изменения, выработавшиеся у рыб в результате такой «тренировки», передавались потомству. В начале XX века, однако, стало ясно, что подобные изменения, выработавшиеся в течение жизни, наследоваться не могут. Следовательно, если дарвиновские представления об эволюции путем постепенных изменений верны, движущей силой процесса мог быть только естественный отбор врожденных наследственных отклонений. Иными словами, предки камбал, у которых «нижний» глаз от рождения был чуть-чуть сдвинут к макушке, должны были оставлять больше потомства, чем те, у которых череп был строго симметричен, а «нижний» глаз смотрел прямо в песок.
Это предположение многим казалось неправдоподобным. Какое преимущество могло дать предкам камбал небольшое смещение глаза, если он все равно оставался на нижней стороне головы и, следовательно, не мог ничего видеть, кроме песка? Противники дарвинизма издавна утверждали, что гипотетические переходные формы между «обычными» рыбами и камбалами — формы, у которых череп уже асимметричен, но глаза еще находятся по разные стороны головы, — не могли возникнуть под действием отбора, да и вообще не могли существовать. Такое строение, говорили они, было бы бессмысленным, неадаптивным.
Подобные рассуждения использовались не только креационистами, но и добросовестными учеными, которые пытались найти выход из кажущегося тупика, предлагая альтернативные эволюционные теории. В частности, эволюция камбал была любимым примером Рихарда Гольдшмидта (см. также Richard Goldschmidt), который выдвинул идею скачкообразной эволюции (теория макромутаций или «перспективных монстров» («Hopeful Monster» hypothesis). Гольдшмидт полагал, что камбалы появились в результате одной крупной мутации, которая сразу перенесла глаз на другую сторону головы. Получившийся «монстр» оказался «перспективным», он сразу получил адаптивное преимущество, и его потомки размножились в морях.
Идеи Гольдшмидта в настоящее время не пользуются большой популярностью, хотя никто не отрицает, что крупные мутации время от времени происходят. Но вот вероятность того, что такая мутация окажется полезной, по-видимому, чрезвычайно мала. Гораздо более типичной является «дарвиновская», то есть более или менее постепенная эволюция. Но в этом случае должны были существовать переходные формы между «обычными» рыбами и камбалами, а найти их до сих пор не удавалось.
Самыми примитивными представителями камбалообразных до сих пор считались псеттоды (род Psettodes), у которых «смещенный» глаз находится на верхнем ребре головы. Но все-таки этот глаз уже не зарывается в песок, когда рыба лежит на дне.
|

И вот наконец долгожданное «недостающее звено» между нормальными рыбами и камбалообразными найдено. Честь открытия принадлежит Мэтту Фридману (Matt Friedman) из Чикагского университета, опубликовавшему в последнем номере журнала Nature статью с описанием двух ископаемых рыб из отложений ипрского и лютетского веков (55,8–40,4 млн лет назад, эоценовая эпоха) Франции и Северной Италии.
Первая рыба, получившая название Heteronectes chaneti, ранее не была известна науке. Вторая (род Amphistium с двумя видами: paradoxum и altum) уже была известна, но Фридману впервые удалось точно реконструировать строение ее черепа при помощи рентгеновской томографии.
Эти две рыбы имеют то самое строение черепа, которое противники Дарвина считали «невозможным»: череп резко асимметричен, одна глазница уже сместилась наверх, но оба глаза еще находятся по разные стороны головы. Это явно не ошибка и не результат посмертных изменений ископаемых костей. Среди фактов, подтверждающих это, можно упомянуть асимметричное строение лобных костей у Heteronectes. Правая лобная кость крупная, прямоугольной формы; левая же превратилась в узкую изогнутую пластинку, ограничивающую верхний край сместившейся вверх глазницы. Род Amphistium известен по множеству экземпляров, и строение костей черепа у всех более или менее одинаково. Во всех случаях череп асимметричен, одна из глазниц смещена вверх, но остается на «своей» стороне черепа.
Все найденные экземпляры Heteronectes и Amphistium как минимум на порядок крупнее мальков современных камбалообразных на той стадии, когда глаза у них оказываются на одной стороне головы. Даже у примитивного (или, как сейчас принято говорить, «морфологически консервативного») псеттода глаз оказывается на верхнем ребре головы уже при длине тела 13 мм, тогда как длина типового экземпляра Heteronectes — 142 мм. Следовательно, не может быть и речи о том, что найденные переходные формы представляют собой мальков, у которых миграция глазницы еще не завершилась. Об этом же свидетельствует и целый ряд других признаков, таких как полное окостенение черепа и отсутствие возрастных изменений в положении глазницы у Amphistium в процессе роста от 103 до 200 мм. Несомненно, эти рыбы — взрослые, и глазницы у них находятся в своем окончательном положении.
Не только строение черепа, но и другие морфологические признаки Heteronectes и Amphistium свидетельствуют о том, что это самые примитивные из известных на сегодняшний день камбалообразных. В их скелете имеются некоторые архаичные признаки, характерные для предков камбалообразных — древних представителей надотряда Perciformes. Некоторые из этих признаков не сохранились ни у кого из современных камбалообразных, другие сохранились только у псеттода (например, шипы в спинном и анальном плавниках).
Любопытно, что среди экземпляров Amphistium есть как «левые», так и «правые» особи. То же самое наблюдается у псеттода, однако у всех более продвинутых камбалообразных такие вариации крайне редки: каждый вид обычно представлен или только «левыми», или только «правыми» рыбами.
Находки убедительно показали, что становление камбалообразных все-таки шло «по Дарвину» — постепенно, а не «по Гольдшмидту» — одним скачком. Следовательно, постепенные преобразования черепа в индивидуальном развитии современных камбалообразных действительно воспроизводят ход их эволюции, что соответствует «биогенетическому закону» Геккеля (см. также Recapitulation theory); этот закон в своей жесткой и абсолютизированной форме давно отвергнут наукой, но в смягченном варианте часто оказывается вполне справедлив.
Ископаемые рыбы, описанные Фридманом, опровергают утверждение о «невозможности» и «нежизнеспособности» переходных форм между камбалами и нормальными рыбами с симметричным черепом. Однако хотелось бы еще и понять, как же все-таки древние камбалообразные использовали свой нижний глаз, который должен был, по идее, глядеть прямо в песок. Фридман отмечает, что подсказку можно найти в поведении некоторых современных камбалообразных, которые время от времени приподнимают свое тело над поверхностью дна, опираясь на лучи спинного и анального плавников. Heteronectes и Amphistium тоже могли «отжиматься» таким образом, поскольку спинной и анальный плавники у них были очень мощные. Приподнимая голову над песком, эти рыбы могли своим смещенным нижним глазом осматривать поверхность дна перед собой в поисках добычи.
|

Необходимо отметить, что Heteronectes и Amphistium не были прямыми предками современных камбалообразных. Они были «всего-навсего» очень близкими родственниками этих предков, точнее — малоизменившимися потомками тех рыб, что дали начало всем камбалообразным, современным и вымершим. Впрочем, то же самое можно сказать практически о любой ископаемой переходной форме: вероятность найти в палеонтологической летописи чьего-нибудь прямого предка очень мала, и по умолчанию среди биологов принято такую возможность вообще не рассматривать. Поэтому любая переходная форма автоматически интерпретируется как «боковая веточка», а не «участок ствола».
По-видимому, первые камбалообразные появились еще в палеоцене (65,6–55,8 млн лет назад) и очень быстро разделились на множество эволюционных линий. В эоцене, когда жили Heteronectes и Amphistium, уже существовали более продвинутые камбалообразные с глазами на одной стороне головы. Таким образом, удивительные рыбы, описанные Фридманом, уже в эоцене были своего рода живыми ископаемыми, то есть мало изменившимися («морфологически консервативными») потомками общего предка группы. В эоценовых морях они смотрелись примерно так же, как в современных смотрится псеттод с глазом на макушке.

")