|
Многие теоретики признают возможность действия отбора не только на уровне генов и особей, но и на более высоких уровнях, в том числе на уровне видов. Межвидовой отбор может иметь место, если какие-то наследственные признаки, передающиеся от родительских видов к дочерним, существенно влияют на темп диверсификации (r), который представляет собой разность скоростей (или вероятностей) появления видов (?) и их вымирания (?).
Межвидовой отбор, разумеется, работает гораздо медленнее, а значит, и не так эффективен, как отбор на уровне особей. Если между двумя типами отбора возникает конфликт (например, если какой-то признак вреден для особей, но полезен для ускоренной диверсификации видов), «победа» в подавляющем большинстве случаев будет на стороне индивидуального отбора. Вредный для особей ген (или, что в большинстве случаев то же самое, аллель, хуже справляющийся с задачей собственного размножения по сравнению с конкурирующим аллелем) просто будет элиминирован из генофонда популяции благодаря обычному, индивидуальному отбору — и межвидовой отбор решительно ничего не сможет с этим поделать. Если данная эволюционная линия в итоге вымрет — что ж, тем хуже для нее. Отбор не заглядывает в будущее, он отслеживает только сиюминутные выигрыши и проигрыши для размножающихся объектов (подробнее см. в обзоре «Доказательства эволюции»).
Могут быть, однако, и такие ситуации, в которых межвидовой отбор, несмотря на всю свою медленность и низкую эффективность, всё-таки способен повлиять на макроэволюционные процессы. Например, если признак, поддерживаемый межвидовым отбором, с точки зрения генов и особей является нейтральным или если частота мутаций, ведущих к исчезновению данного признака, очень низка (сопоставима с темпами появления и вымирания видов). Однако до сих пор известно очень мало конкретных фактов, указывающих на действенность межвидового отбора (Jablonski, 2008. Species Selection: Theory and Data; Rabosky & McCune, 2010. Reinventing species selection with molecular phylogenies).
|
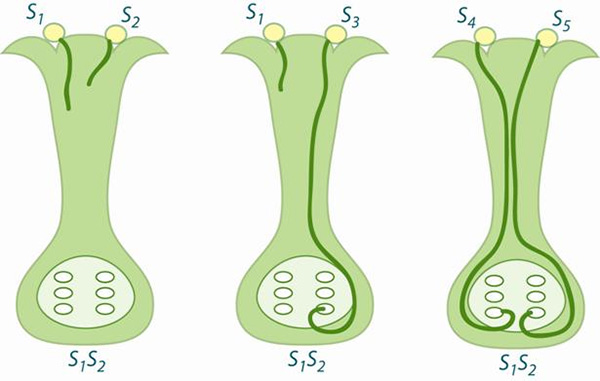
Проблема тут в том, что, хотя разные группы организмов могут сильно различаться по темпам появления и вымирания видов, эти различия, как правило, трудно увязать с какими-то конкретными признаками (морфологическими, физиологическими или поведенческими). Американские и британские биологи выбрали для проверки предположения о действенности межвидового отбора на редкость удобный объект — семейство пасленовых, и очень подходящий признак — самонесовместимость. Удобность объекта обусловлена огромным видовым разнообразием пасленовых и их хорошей изученностью, в том числе на генетическом уровне. Самонесовместимость, или отторжение родственной пыльцы (рис. 1), интересна тем, что, во-первых, этот признак, исходя из общих соображений, вполне может влиять на темпы видообразования и вымирания, во-вторых — и это главное — он распространен среди видов пасленовых достаточно хаотично. Во многих родах пасленовых одни виды имеют систему самонесовместимости, тогда как другие, в том числе близкородственные, виды такой системы не имеют. При этом наличие или отсутствие самонесовместимости практически не коррелирует с другими признаками этих растений. Это дает основания надеяться, что если удастся выявить корреляцию между самонесовместимостью и темпами диверсификации, то эта корреляция будет отражать причинную связь.
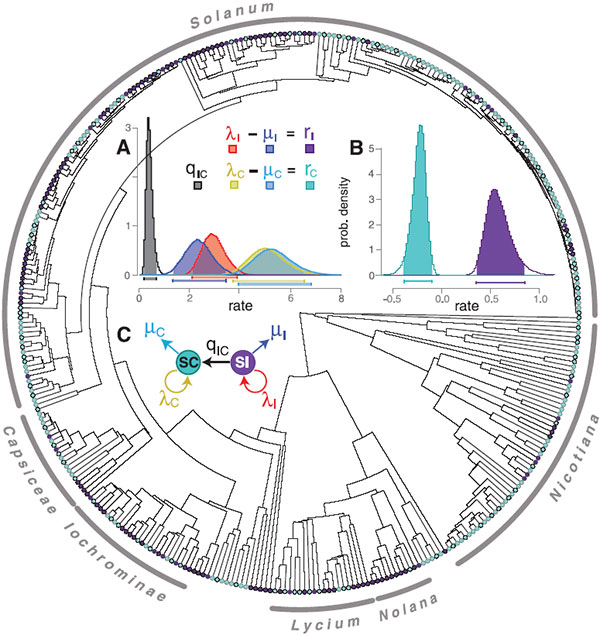
В семействе пасленовых около 2700 видов, из которых примерно 41% имеют систему самонесовместимости, 57% ее не имеют, а 2% видов — двудомные, то есть имеющие отдельные мужские и женские растения, так что для них проблема самооплодотворения не актуальна. Авторы построили филогенетическое (эволюционное) дерево для 356 видов пасленовых, по которым имеются необходимые молекулярные данные (дерево строилось по последовательностям двух ядерных генов и четырех пластидных) и для которых точно установлено наличие или отсутствие механизма самонесовместимости.
Анализ получившегося дерева показал (впрочем, это было понятно и раньше), что самонесовместимость унаследована пасленовыми от общего предка и с тех пор многократно утрачивалась в разных эволюционных линиях. Утратить эту систему легко, а восстановить обратно — трудно, потому что это сложный молекулярный комплекс, в котором участвует множество специализированных белков. Судя по всему, в эволюции пасленовых почти или вовсе не было случаев восстановления самонесовместимости после ее утраты.
Почему самонесовместимость часто утрачивается, более или менее понятно. Переход к самооплодотворению дает немедленное преимущество в эффективности распространения собственных генов (см.: Опыты на червях доказали, что самцы — вещь полезная, «Элементы», 23.10.2009); к тому же самооплодотворение может давать адаптивное преимущество, когда с доставкой пыльцы от неродственных особей возникают трудности — например, из-за большой разреженности популяции (см.: Чтобы превратить самок в гермафродитов, достаточно двух мутаций, «Элементы», 16.11.2009). Непонятно другое: если этот признак часто утрачивается и почти никогда не восстанавливается, почему до сих пор сохранилось так много видов, обладающих системой самонесовместимости?
Чтобы ответить на этот вопрос, авторы проанализировали филогенетическое дерево пасленовых при помощи новой методики, которая называется BiSSE (binary state speciation and extinction model); см.: Maddison et al., 2007. Estimating a Binary Character's Effect on Speciation and Extinction. Этот метод предназначен как раз для анализа зависимости скоростей появления и вымирания видов от какого-нибудь бинарного (то есть принимающего одно из двух значений) признака, такого как наличие или отсутствие самонесовместимости. Метод позволяет подобрать шесть наиболее подходящих к данному дереву параметров: ?1 и ?2 — средние скорости видообразования для видов с двумя альтернативными состояниями признака, ?1 и ?2 — скорости вымирания, q12 и q21 — вероятности перехода признака из состояния 1 в 2 и обратно. В данном случае вероятность перехода от отсутствия самонесовместимости к ее наличию считалась равной нулю.
Расчеты показали, что темп видообразования у видов, практикующих самоопыление, значительно выше, чем у самонесовместимых. Однако темп вымирания у них еще выше, так что итоговая скорость диверсификации (r = ? – ?) оказывается выше у видов, обладающих системой самонесовместимости. Несмотря на то, что совокупность самоопыляющихся видов постоянно пополняется за счет превращения самонесовместимых видов в самоопыляющиеся, а обратное превращение «запрещено», численность самонесовместимых видов не снижается до нуля, а остается на постоянном уровне (порядка 30–40%), потому что такие виды эффективнее «размножаются», передавая своим видам-потомкам самонесовместимость по наследству. Это и есть межвидовой отбор в действии: именно благодаря межвидовому отбору самонесовместимость до сих пор не исчезла у пасленовых.
Повышенная скорость видообразования у растений, способных к самоопылению, очевидно, связана с тем, что у них не так остро стоит проблема «размывания» полезных комбинаций аллелей, сложившихся в ходе приспособления к местным условиям. Одно-единственное растение, попавшее в необычные условия, способно дать начало новому виду. Почему они чаще вымирают, тоже в общем-то понятно: у них должны быстрее накапливаться вредные мутации и реже фиксироваться мутации полезные (подробнее о пользе перекрестного оплодотворения см. в заметке Опыты на червях доказали, что самцы — вещь полезная, «Элементы», 23.10.2009).
Данная работа показала, что межвидовой отбор способен оказывать заметное влияние на макроэволюцию. Он может обеспечить длительное сохранение сложного признака, который в каждой отдельно взятой эволюционной линии имеет тенденцию исчезать и почти никогда не появляется вновь. Но нужно помнить, что медлительный и малоэффективный межвидовой отбор, конечно, не в состоянии создать такой признак «с нуля»: подобным творческим потенциалом обладает только отбор на более низких уровнях (в первую очередь на уровне генов и особей).

")