|
Бактерия вольбахия живет в цитоплазме клеток многих насекомых, паукообразных, ракообразных и круглых червей. Она лишь изредка передается горизонтально от одного хозяина к другому, а главный путь ее распространения — от матери к детям с цитоплазмой яйцеклетки. Иными словами, гены вольбахии распространяются в популяции хозяев практически так же, как гены митохондрий. Самцы с точки зрения вольбахии являются «тупиком»: бактерии, попавшие в самца, обречены погибнуть вместе с ним и не могут перейти в следующее поколение хозяев. Поэтому между генами вольбахии и ядерными генами хозяина возникает конфликт: первые заинтересованы в том, чтобы зараженная самка производила на свет только дочерей, а интересы ядерных генов, как правило, состоят в том, чтобы соотношение полов у потомства было равным. Тем самым создаются предпосылки для эволюционной «гонки вооружений». Можно ожидать, что в геноме вольбахии будут закрепляться мутации, способствующие увеличению доли дочерей в потомстве зараженной самки. В качестве «ответной меры» в ядерном геноме хозяина могут накапливаться мутации с обратным фенотипическим эффектом, то есть способствующие увеличению доли сыновей.
Митохондрии находятся в таком же положении, что и вольбахия, но обычно ничего не могут с этим поделать, потому что в митохондриальном геноме очень мало белок-кодирующих генов (у большинства животных — менее двух десятков). Большинство генов, необходимых для жизни и размножения митохондрий, находятся в ядре. Из-за этого способности митохондрий к эгоистической эволюции крайне ограничены. Впрочем, у растений митохондриальные геномы значительно больше, при этом известны эгоистические митохондрии-мутанты, вызывающие гибель мужских половых клеток у некоторых растений (это явление называется «цитоплазматической мужской стерильностью»). Вольбахия сохранила свыше 1000 собственных генов, что позволило ей добиться куда более впечатляющих успехов в борьбе с мужским полом (см. ссылки в заметке Животные обмениваются генами с паразитическими бактериями, «Элементы», 05.09.2007).
Вольбахия использует для уменьшения числа самцов различные методы — от грубого «андроцида» (избирательного умерщвления зараженного потомства мужского пола) до изощренной «феминизации» (превращения самца в самку). К перепончатокрылым насекомым вольбахия нашла особый подход, связанный с необычным способом наследования пола в этом отряде. У перепончатокрылых самки диплоидны и развиваются из оплодотворенных яиц, то есть имеют и мать, и отца. Самцы гаплоидны, развиваются из неоплодотворенных яиц и отца не имеют. Самка после спаривания сохраняет полученную сперму в специальном резервуаре и в дальнейшем сама может решать при откладке очередного яйца, оплодотворить ли его (и тогда будет дочка), или оставить неоплодотворенным (тогда будет сын).
Вольбахии, паразитирующие в клетках перепончатокрылых, научились блокировать расхождение хромосом во время первого или второго митотического деления неоплодотворенной яйцеклетки. В результате исходно гаплоидный зародыш, который должен был бы стать самцом, оказывается диплоидным (и при этом гомозиготным по всем локусам) и развивается в самку.
Эта манипуляция называется «индукцией партеногенеза» (потому что зараженные самки начинают производить на свет дочерей партеногенетическим путем), а бактерии, научившиеся этому трюку, обозначаются буквами PI (parthenogenesis inducing). PI-вольбахии очень быстро распространяются в популяциях хозяев за счет того, что зараженные ими самки начинают производить вдвое больше дочерей, чем незараженные. Фактически PI-вольбахии позволяют зараженным самкам не платить так называемую «двойную цену пола», о которой говорилось в заметке Опыты на червях доказали, что самцы — вещь полезная («Элементы», 23.10.2009). Поскольку большинство или все дочери зараженной самки тоже оказываются зараженными, инфекция вскоре может достичь 100-процентной частоты в популяции.
Действительно, большинство популяций наездников либо заражены PI-вольбахией поголовно, либо не заражены вовсе. Это значит, что если уж инфекция проникла в популяцию и не была элиминирована сразу, то она, скорее всего, достигнет 100-процентной частоты.
Лишь у некоторых видов наездников из рода Trichogramma встречаются смешанные популяции, включающие как зараженных, так и здоровых насекомых. Эти виды, вероятно, научились как-то противодействовать распространению инфекции. В одном случае (у вида T. kaykai) известен механизм этого противодействия. В генофонде этого вида имеется особая «эгоистическая» хромосома (ее обозначают буквами PSR — paternal sex ratio), передающаяся строго по мужской линии. Когда эта хромосома при оплодотворении попадает в зиготу, она уничтожает все отцовские хромосомы, кроме самой себя. В результате яйцо оказывается де-факто неоплодотворенным. Поэтому из него развивается не самка, а гаплоидный самец — обладатель PSR-хромосомы. Как видим, PSR-хромосома и PI-вольбахия совершают действия, прямо противоположные по своему смыслу: первая превращает диплоидный эмбрион в гаплоидный, то есть делает из самок самцов, а вторая гаплоидный эмбрион делает диплоидным, превращая исходно мужской зародыш в женский. Это типичный результат эволюционной «гонки вооружений». В результате в популяциях, где есть PSR-хромосома, инфицированность PI-вольбахией поддерживается на постоянном невысоком уровне (около 10%).
Изучение смешанных популяций Trichogramma показало, что PI-вольбахия вовсе не препятствует нормальному половому размножению насекомых. Зараженные самки нормальным образом спариваются с самцами, и, если самец не является носителем PSR-хромосомы, из оплодотворенных яиц развиваются нормальные самки. Эти самки тоже оказываются заражены вольбахией, поэтому их существование нисколько не ущемляет интересы паразита. В итоге зараженные самки Trichogramma производят на свет дочерей двух сортов: «нормальных», получившихся из оплодотворенных яиц, и «искусственно созданных» вольбахией из неоплодотворенных яиц, которые в норме должны были бы дать самцов. Эти две разновидности самок можно отличить друг от друга по уровню гомозиготности: самки, развившиеся из неоплодотворенных яиц, имеют два идентичных хромосомных набора, то есть гомозиготны по всем локусам.
Все факты, о которых шла речь до сих пор, на сегодняшний день вполне объяснимы и понятны. Но у многих видов зараженных вольбахией наездников есть одна странная черта, которую до сих пор не удавалось внятно объяснить. Практически во всех популяциях, поголовно зараженных PI-вольбахией, самки (но не самцы!) полностью утратили способность к нормальному половому размножению. Даже если вылечить этих самок от вольбахии (это нетрудно сделать при помощи антибиотиков), они могут спариваться с самцами, но всё равно продолжают откладывать исключительно неоплодотворенные яйца. Теперь, без вмешательства вольбахии, из этих яиц развиваются уже не самки, а самцы — вполне нормальные самцы, способные оплодотворять самок из незараженных популяций. Эти самцы могут спариваться и с вылеченными самками из зараженной популяции, но потомства от таких спариваний не бывает, самки не используют полученную сперму и по-прежнему откладывают только неоплодотворенные яйца, из которых выводятся только самцы. Таким образом, получается, что в поголовно зараженных популяциях нормальная репродукция без вольбахии невозможна. Без «помощи» бактерии в таких популяциях могут рождаться только самцы, что, конечно же, приведет популяцию к полному вымиранию уже во втором поколении.
Почему самки в зараженных популяциях утратили способность к половому размножению, тогда как самцы ее сохранили? Над этой загадкой ученые долго ломали головы. Проще всего объяснить утрату полового размножения тем, что в полностью зараженной популяции самцов вообще нет, спариваться не с кем, всё размножение идет путем партеногенеза, и поэтому мутации, нарушающие работу генов, необходимых для нормального полового размножения, перестают проявляться в фенотипе и отсеиваться отбором. Накопление таких мутаций со временем приводит к необратимой утрате способности к половому размножению.
Объяснение в принципе неплохое, но оно не дает ответа на вопрос, почему всё это происходит только с самками, а самцы, которых в зараженной популяции вообще не было до применения антибиотика, после лечения «воскресают» здоровенькими и вполне плодовитыми? Ведь вредные мутации, нарушающие мужскую репродуктивную функцию, в зараженной популяции тоже не отсеивались отбором! Очевидно, потеря самками способности к половому размножению должна была давать этим самкам какое-то преимущество. Можно было бы предположить, что это преимущество «экономического» характера: перестав тратить ресурсы на ставшую ненужной функцию (например, на развитие сперматеки и других органов, необходимых для спаривания и оплодотворения), самки могли бы направить больше ресурсов на что-то другое, например на производство дополнительных яиц. Однако факты не подтверждают это предположение: нет данных, которые указывали бы на повышенную плодовитость самок, утративших половое размножение.
Правдоподобное решение этой непростой задачи нашли генетики из США и Франции, опубликовавшие свои результаты в журнале BMC Evolutionary Biology. Их главный вывод состоит в том, что утрата самками способности к половому размножению при определенных условиях возникает как вполне предсказуемый и даже неизбежный побочный результат эволюционной «гонки вооружений», порожденной конфликтом интересов генов PI-вольбахии и насекомого.
Авторы решали эту теоретическую задачу при помощи моделирования, причем параллельно использовалось два подхода: аналитический (то есть выведение устрашающих формул) и имитационный (когда всю ситуацию моделируют на компьютере, вводят те или иные исходные параметры, дают виртуальной популяции спокойно жить и эволюционировать в течение скольких-то поколений, а затем смотрят, что получилось). Оба подхода привели авторов к одним и тем же выводам, которые состоят в следующем.
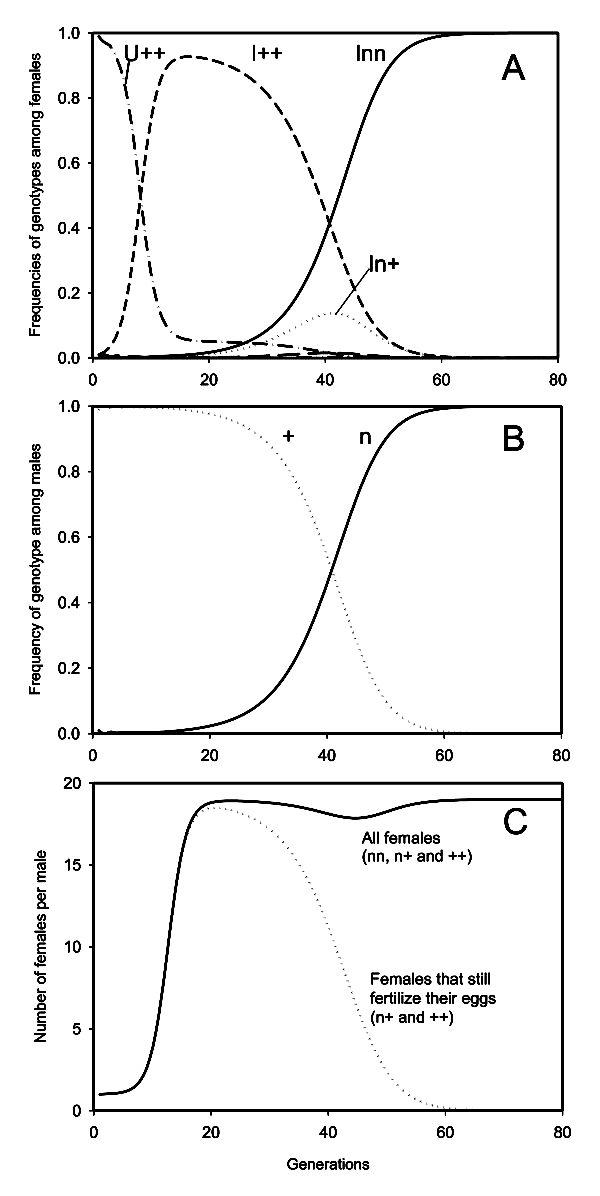
Если PI-вольбахия, впервые попавшая в незараженную популяцию, сразу будет обладать способностью проникать в 100% яиц, отложенных зараженной самкой, и если инфекция не приводит к сильному снижению жизнеспособности и плодовитости, то очень быстро зараженность популяции станет 100-процентной (по причинам, рассмотренным выше). Самцы исчезнут, половое размножение прекратится, и больше ничего особо интересного не произойдет: эволюционная «гонка вооружений» просто не успеет развернуться.
Такой сценарий, однако, маловероятен. Как показывают эксперименты, при заражении новой популяции вольбахиям практически никогда не удается сразу обеспечить себе гарантированное проникновение в каждое яйцо зараженной самки. Бактериям нужно значимое по эволюционным меркам время, чтобы добиться такой эффективности наследования.
Если же заражаемость яиц поначалу не стопроцентная, то инфекция будет распространяться медленнее, и в популяции довольно долго будет сохраняться некоторое количество самцов и незараженных самок. В такой ситуации, как видно из представленных авторами моделей, любая мутация, заставляющая самок реже использовать сперму и откладывать меньше оплодотворенных яиц, будет поддержана отбором и довольно быстро зафиксируется, то есть достигнет 100-процентной частоты в популяции наездников. Этот результат не зависит от того, насколько силен эффект мутации. Мутация может лишь ненамного уменьшить долю оплодотворяемых яиц, а может сразу свести ее к нулю (авторы называют такие мутации «мутациями функциональной девственности») — всё равно отбор такую мутацию поддержит и зафиксирует. Отбор будет закреплять эти мутации, одну за другой, до тех пор, пока все самки в популяции полностью не прекратят использовать сперму для оплодотворения яиц и не станут производить потомство только партеногенетическим путем. Произойдет то, что и наблюдается у многих наездников — полная и необратимая потеря самками способности к половому размножению, не восстанавливающаяся даже после утраты PI-вольбахии. К этому результату система неизбежно приходит при любых значениях входных параметров, с одной-единственной оговоркой: зараженные самки должны производить в среднем больше дочерей, чем незараженные (а это условие в реальности практически всегда выполняется).
Почему же отбор так решительно поддерживает любые мутации, снижающие долю оплодотворенных яиц? Это происходит благодаря последовательному действию двух механизмов. Сначала, пока в популяции еще много незараженных особей, будет действовать механизм, открытый еще в 1930-е годы великим эволюционистом-теоретиком Рональдом Фишером. Это тот самый механизм, который обеспечивает у большинства животных равное соотношение полов в популяциях (см.: Почему соотношение полов почти всегда 1:1). Распространение PI-вольбахии приводит к снижению доли самцов. Самцы становятся «дефицитным» полом: на каждого самца теперь приходится более одной самки. Следовательно, в среднем каждый самец будет оставлять больше потомков, чем средняя самка. С эволюционной точки зрения это означает, что быть самцом теперь «выгоднее», чем самкой. Следовательно, самке становится выгоднее рожать сыновей, чем дочерей, потому что это повышает эффективность тиражирования ее генов в следующих поколениях (проще говоря, такая самка оставит больше внуков). Таким образом, отбор будет поддерживать любые мутации, способствующие рождению сыновей, а не дочерей. Для самок перепончатокрылых, как мы знаем, самый простой и естественный способ рожать больше сыновей — это не оплодотворять свои яйца.
В результате в популяции происходит параллельное распространение и PI-вольбахии, и мутаций, снижающих частоту оплодотворения. В конце концов «фишеровский» механизм перестает работать, потому что в популяции остается слишком мало самок, способных к оплодотворению. В такой ситуации быть самцом уже не выгодно, даже если самцы — большая редкость.
Самый интересный и неожиданный вывод, сделанный авторами, состоит в том, что задолго до того, как перестанет работать «фишеровский» механизм, распространение «генов отказа от оплодотворения» начнет подстегиваться другим эффектом, оценить действенность которого без имитационного моделирования было бы трудно. Теперь, когда самцы уже не имеют адаптивного преимущества перед самками, распространение мутантных генов будет продолжаться благодаря тому, что подавляющее большинство оставшихся в популяции самцов на этом этапе являются потомками мутантных самок и сами несут в своем гаплоидном геноме мутантные гены, снижающие частоту оплодотворения. Ведь мы помним, что мутации, о которых идет речь, увеличивают производство сыновей. Мутантные самки производят больше сыновей, передавая им «гены отказа от оплодотворения», а сыновья, в свою очередь, передают их всем своим дочерям. В результате доля самок — обладательниц этих генов будет продолжать расти, хотя это и не приносит им больше «фишеровской» выгоды в виде дополнительных внуков.
Таким образом, самцы становятся главными распространителями генов, которые в краткосрочной перспективе способствуют увеличению численности самцов, а в долгосрочной делают их вовсе ненужными.
Когда частота «генов функциональной девственности» приблизится к 100%, рожать самцов станет вовсе бессмысленно — ведь самцам теперь некого оплодотворять, и у самца не будет шансов передать свои гены потомкам. Теперь выгодно рожать только самок, но наши наездники уже разучились это делать без помощи вольбахии! Таким образом, размножение насекомых оказывается в полной зависимости от паразитической бактерии. Бактерия становится как бы частью репродуктивной системы самок, частью, без которой они не способны произвести на свет ничего, кроме бессмысленных самцов, не способных оставить потомство в этой популяции. Таким образом, бактерия из наглого паразита, бесцеремонно манипулирующего своими хозяевами, формально превращается в полезного симбионта, без которого хозяева теперь не в состоянии обойтись.
|
В этой новой ситуации наездники оказываются заинтересованы в том, чтобы вольбахия попадала в каждое яйцо (раньше в этом была заинтересована только сама вольбахия). Пустив вольбахию в каждое яйцо, наездники окончательно избавятся от ставших никому не нужными самцов. Любая мутация, повышающая заражаемость яиц, произошедшая в геноме вольбахии или наездника, будет немедленно поддержана отбором. В результате получим картину, характерную для большинства зараженных популяций наездников: зараженность полная, самцов нет, передача вольбахии по наследству близка к 100%, все самки лишены способности к половому размножению, а самцы, которых можно «вызвать из небытия», вылечив самку антибиотиком и получив от нее потомство, — вполне способны оплодотворить самку из другой, незараженной популяции того же вида наездников.
Данная работа интересна, во-первых, как пример изящного решения довольно сложной задачи по эволюционной генетике. Во-вторых, она показывает, что эволюционная гонка вооружений, порождаемая конфликтом между генами хозяина и внутриклеточного паразита, может в итоге превратить паразита в полезного симбионта и чуть ли не в неотъемлемую часть репродуктивной системы хозяина. Долгая предварительная история мирного сосуществования и взаимовыгодного сотрудничества для этого вовсе не обязательна.

")