Анализ большого массива палеонтологических данных показал, что роды морских животных, приуроченные к палеосообществам с высоким родовым разнообразием, существовали в среднем дольше и вымирали реже по сравнению с родами, жившими в простых сообществах. Этот факт подтверждает гипотезу, согласно которой высокое биоразнообразие повышает устойчивость сообществ. Споры вокруг этой гипотезы идут уже полвека, однако до сих пор все аргументы «за» и «против» экологи получали только из анализа современных экосистем.
|

Например, если в сообществе много видов, то они, скорее всего, будут в среднем иметь более узкие ниши: каждый хищник будет охотиться на немногие виды жертв, каждое травоядное — питаться ограниченным кругом растений и т. д. Если численность какого-нибудь вида в таком сообществе начнет снижаться, это, скорее всего, приведет к тому, что условия для жизни этого вида станут более благоприятными: пищи станет больше, специализированных хищников и паразитов — меньше. Вид, начавший вымирать, получит шанс восстановиться.
Если же в сообществе, к примеру, только один вид хищников, охотящийся на многих жертв, то снижение численности одной из жертв не скажется на численности хищников, и они могут спокойно «доесть» вымирающий вид. Впрочем, можно придумать и множество других умозрительных моделей, в которых все будет по-другому, и, главное, гораздо сложнее и неоднозначнее (см., например: Правильно выбирая друзей, можно спастись от конкурентов, «Элементы», 28.04.2009).
Многолетнее изучение различных современных экосистем — естественных и искусственных — так и не дало простого и четкого ответа на вопрос о соотношении разнообразия и устойчивости. По-видимому, это соотношение действительно весьма неоднозначно. В одних ситуациях разнообразие способствует устойчивости, в других — нет, и какая из тенденций преобладает в глобальном масштабе — сказать трудно.
Для разрешения этой дилеммы было бы очень полезно привлечь данные по минувшим геологическим эпохам, однако до сих пор сделать это было трудно из-за отсутствия достаточно полных глобальных сводок по древним сообществам (точнее, по комплексам ископаемых, на основе которых можно реконструировать свойства давно исчезнувших сообществ). Сейчас такие сводки наконец начали появляться. Самая большая и подробная из них — международная база данных The Paleobiology Database (PBDB). На сегодняшний день в нее внесено около 80 000 палеонтологических коллекций. Под «коллекцией» в PBDB понимается комплекс ископаемых, найденных в одном месте и в одном геологическом слое. В большинстве случаев такая коллекция представляет собой более или менее репрезентативную выборку из реального палеосообщества (или из нескольких сообществ, существовавших в данном районе на протяжении некоторого отрезка времени, не слишком большого по геологическим меркам). По среднему числу родов в коллекциях можно оценивать уровень разнообразия древних сообществ — конечно, очень приблизительно, с множеством поправок и оговорок и только при наличии данных по очень большому числу коллекций.
Устойчивость палеосообществ измерить напрямую затруднительно, но можно использовать параметр, тесно с ней связанный и достаточно точно определяемый по палеонтологической летописи, — продолжительность существования видов или родов, входящих в состав этих палеосообществ. Очевидно, что роды, приуроченные к устойчивым сообществам, должны, при прочих равных, вымирать реже и, следовательно, жить дольше по сравнению с родами, приуроченными к неустойчивым сообществам.
Эти соображения легли в основу исследования, результаты которого опубликованы в последнем номере журнала Paleobiology. Работа выполнена автором этих строк. Были использованы данные из PBDB по всем морским животным фанерозойского эона (последние 542 млн лет земной истории). Как выяснилось, между продолжительностью L существования рода и средним родовым богатством D коллекций, в которых есть представители этого рода, существует положительная корреляция. Иными словами, роды, жившие в богатых и разнообразных сообществах, существовали дольше, чем роды, приуроченные к бедным сообществам.
|
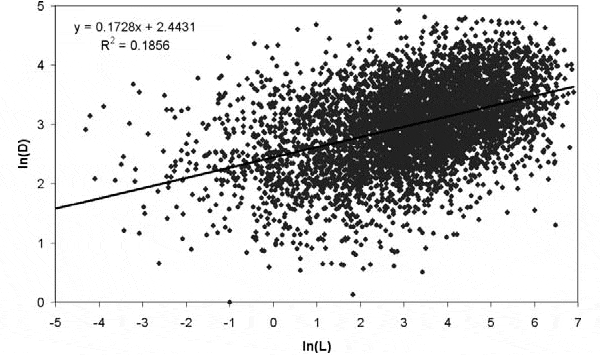
Однако в подобных исследованиях найти искомую закономерность — даже не полдела, а лишь самое начало работы. Большую часть статьи пришлось посвятить рассмотрению возможных альтернативных объяснений найденной зависимости. Ведь положительная корреляция вовсе не обязательно означает, что между двумя величинами существует причинно-следственная связь (то есть, в данном случае, что высокое разнообразие способствует устойчивости). Корреляция может быть вызвана «посторонними» факторами, которые влияют на L и D сходным образом.
Например, можно предположить, что вблизи экватора, где сообщества более разнообразны, роды вымирают реже, чем в высоких широтах — не потому, что живут в разнообразных сообществах, а просто потому, что теплый климат, возможно, снижает вероятность вымирания. Чтобы это проверить, были проанализированы по отдельности коллекции из разных широтных зон. Оказалось, что в пределах отдельных зон (по 10° палеошироты каждая) положительная корреляция всё равно сохраняется. Значит, дело не в широтном градиенте. Аналогичным образом были проверены и другие гипотезы о возможном влиянии различных факторов, в том числе глубинного градиента (рассматривались по отдельности глубоководные и мелководные сообщества) и степени литификации пород. В литифицированных, то есть окаменевших, породах ископаемые сохраняются хуже, чем в сыпучих, и поэтому литификация может одновременно снижать как D, так и L, порождая тем самым наблюдаемую положительную корреляцию. Оказалось, что даже с учетом всех этих факторов положительная корреляция между L и D всё равно остается значимой.
В числе прочих было проверено и влияние «временного градиента». Дело в том, что и средняя продолжительность существования родов, и среднее родовое богатство морских сообществ в течение фанерозоя в целом росли. Может быть, именно этот параллельный рост объясняет обнаруженную корреляцию? Чтобы проверить это, пришлось рассмотреть по отдельности коллекции, приуроченные к отдельным временным интервалам, в том числе таким, в течение которых ни D, ни L не росли. Корреляция эту проверку тоже выдержала.
|
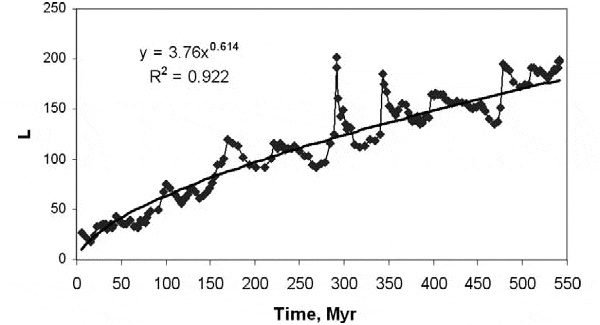
Полученные результаты говорят о том, что в глобальном масштабе влияние разнообразия на устойчивость всё-таки положительное. То есть жизнь в разнообразном сообществе, при прочих равных, снижает вероятность вымирания. Правда, разнообразие сообществ влияет на продолжительность жизни родов не очень сильно. Это видно по низкому значению коэффициента корреляции. Обнаруженная корреляция — слабая, хотя и обладает очень высоким уровнем статистической значимости. Но этого и следовало ожидать, ведь разнообразие сообществ никак не может быть единственным (или основным) фактором, определяющим, сколько просуществует на свете тот или иной род. Продолжительность жизни рода зависит от множества причин, величина D — лишь одна из них. Это примерно как с влиянием курения на продолжительность жизни человека: корреляция очень слабая, но при этом очень значимая.
|
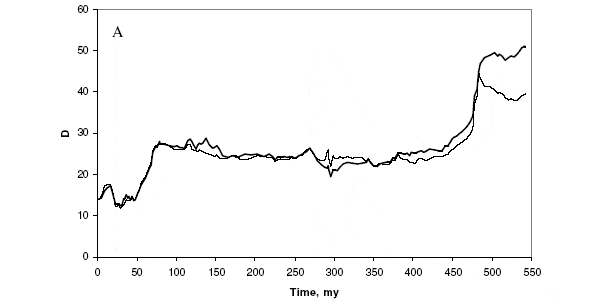
Полученные результаты хорошо согласуются с предложенной нами ранее гиперболической моделью роста биоразнообразия (см.: Биоразнообразие, как и народонаселение, растет по гиперболе; Флора и фауна континентов развивается по гиперболическому закону). Гиперболический характер роста биоразнообразия предполагает, что этот процесс идет под управлением сложных положительных обратных связей. Иными словами, чем выше разнообразие, тем больше создается предпосылок для еще более быстрого его роста. Конечно, такой самоускоряющийся рост не может продолжаться бесконечно, рано или поздно он обязательно начинает замедляться — как это начало происходить с населением мира в 1960-е годы.
Возможны, как минимум, два механизма положительной обратной связи в эволюции биоразнообразия, которые могут объяснить его гиперболический рост. Первый из них проверялся в данной работе и получил подтверждение: рост разнообразия сообществ ведет к росту их устойчивости, а это, в свою очередь, приводит к снижению скорости вымирания родов. В результате общее число родов растет быстрее. Второй механизм основан на том, что новые виды могут создавать ниши для следующего «поколения» новых видов (см.: Цепная реакция видообразования, «Элементы», 11.02.2009). Поэтому, при соблюдении определенных условий, рост разнообразия сообщества может повышать вероятность появления в этом сообществе новых видов. Действенность этого механизма тоже можно оценить на основе данных из PBDB, что мы с коллегами сейчас и пытаемся сделать.

")