|

Массовое вымирание на рубеже триаса и юры входит в пятерку самых крупных экологических кризисов за последние 540 млн лет земной истории (остальные четыре — это массовые вымирания на рубеже ордовика и силура 444 млн лет назад, в конце девона 360 млн лет назад, на рубеже перми и триаса 251 млн лет назад и на рубеже мела и палеогена 65,5 млн лет назад).
Причины глобальных биотических кризисов, вероятнее всего, были разными. Это могли быть падения крупных метеоритов, вызывавших мощные цунами, землетрясения и длительное затемнение атмосферы (см.: неокатастрофизм). Также кризисы могут быть вызваны глобальными изменениями климата — похолоданиями или потеплениями. Распространена и такая точка зрения, что биота, будучи целостной системой, проходит определенные «возрастные» периоды — стадии роста, расцвета и упадка. Достигнув стадии упадка, таксон становится неустойчивым, и любое внешнее воздействие, даже не отраженное в геологических последовательностях, может вызвать вымирание (см.: Взлет и падение видов: новые данные подтверждают старую идею «эволюционного цикла», «Элементы», 20.11.2007). Споры между сторонниками различных гипотез идут и сейчас.
Задавшись вопросом о механизмах биотических кризисов, международная группа палеонтологов из Школы биологии и наук об окружающей среде Дублинского университетского колледжа, факультета наук о Земле Оксфордского университета и отдела палеобиологии Национального музея естественной истории Смитсоновского института (Вашингтон, США) рассмотрела последовательность событий позднетриасового кризиса, который произошел около 200 млн лет назад, вскоре после появления первых динозавров и млекопитающих. Возможно, этот кризис в какой-то мере способствовал тому, что господствующее положение на суше (вплоть до следующего массового вымирания, произошедшего спустя 135 млн лет) заняли именно динозавры, а не зверообразные рептилии и их прямые потомки — млекопитающие.
Одной из наиболее вероятных причин позднетриасового вымирания считается глобальное потепление: средняя температура земной поверхности к концу триаса была на целых 7 градусов выше нынешней. Потепление, в свою очередь, связывают с ростом концентрации углекислого газа в атмосфере. Углекислый газ, возможно, выделялся в больших количествах из мантии, потому что в это время начал раскалываться суперконтинент Пангея, и по краям будущего Атлантического океана образовывались огромные поля вытекающих из недр и застывающих базальтов (см.: Central Atlantic Magmatic Province).
Возможно, биосфера просто «перегрелась», особенно ее тропическая часть. Перегрев тропиков и смена климата с влажного на резко континентальный привели к вымиранию многих видов растений и животных. Для растительного мира в целом этот кризис проявился в основном в смене одних доминирующих групп другими.
Палеоботаники проанализировали в деталях не только общее разнообразие, но и изменения структуры растительных сообществ в ходе кризиса. В качестве модельного примера они использовали шесть последовательных слоев позднетриасовых (рэтских) отложений Гренландии, то есть проследили шесть ступеней биотического кризиса. Как видно на палеокарте, Гренландия в это время находилась на границе влажного и засушливого климата. Растительный мир этой территории был представлен влажными лесами из голосеменных и папоротников.
|
Методика, использованная исследователями, основана на анализе количественного распределения родовых обилий. Рассматриваемые шесть слоев осадочных пород образовались, по-видимому, в результате крупных наводнений, заливавших время от времени низменную лесистую равнину. В породе сохранились многочисленные отпечатки листьев. Авторы подсчитывали по отдельности листья каждого рода и подбирали к получившимся количественным распределениям родовых обилий наиболее подходящую математическую модель (см. подробнее об этой методике в заметке: Великое вымирание 250 миллионов лет назад привело к резкому усложнению морских экосистем, «Элементы», 28.11.2006; заметим, что один из авторов обсуждаемой статьи — Питер Вагнер из Смитсоновского института — является также первым автором статьи про усложнение морских экосистем).
На основе полученных моделей авторы рассчитывали для каждого растительного палеосообщества теоретически ожидаемое число «часто встречающихся» родов. Часто встречающимся считали такой род, один лист которого должен встречаться не реже, чем один раз на каждый миллион листьев. В реальности, конечно, в распоряжении палеонтологов не было никаких миллионов листьев, а всего лишь по несколько сотен из каждого слоя. Однако при помощи чрезвычайно сложных математических методов им удалось «реконструировать» по этой небольшой выборке количественную структуру всего сообщества.
Спрашивается, зачем такие сложности, разве нельзя просто подсчитать количество видов в каждом слое и проследить, как оно менялось со временем? Обычно палеонтологи примерно так и поступают, но такой подход чреват серьезными ошибками. Во-первых, сохранность ископаемых может сильно различаться в разных слоях. Это (а также целый ряд других причин) может приводить к тому, что выборки оказываются несопоставимыми. Для того чтобы сделать их сравнение правомерным, приходится применять сверхсложную статистику (что, собственно, и проделали авторы).
Кроме того, простой подсчет числа таксонов в разных слоях накануне кризисного рубежа, даже с поправками на различия в качестве выборок, часто дает искаженные результаты из-за так называемого эффекта Синьора–Липпса (см.: Signor–Lipps effect). Суть его в том, что из-за неполноты летописи самые последние (по времени существования) представители данного вида или рода почти наверняка не будут обнаружены палеонтологами. Поэтому даже если массовое вымирание на данном кризисном рубеже в действительности было мгновенным, большинство видов «исчезнет» из летописи не на самом рубеже, а за какое-то время до него. В результате мгновенное вымирание будет выглядеть в летописи как постепенное. Важнейшее достоинство той методики, которую применили авторы обсуждаемой статьи, состоит как раз в том, что она нечувствительна к эффекту Синьора–Липпса.
Проведенный анализ показал, что по мере развития кризиса кривые распределения видов менялись: доминантных видов становилось всё больше, а обилие редких видов снижалось. Эта картина свидетельствует об упрощении сообществ и снижении разнообразия. Статистическими методами было показано, что снижение разнообразия происходило не плавно, а скачкообразно. Упрощение сообществ началось более или менее внезапно между 3-м и 4-м слоями (предположительно за 300–500 тыс. лет до конца триаса и начала юры). В этот момент ожидаемое число «часто встречающихся» родов снизилось с 33–40 до 27, в пятом слое оно упало до 20, в шестом — до 15 (см. рис. 3).
|
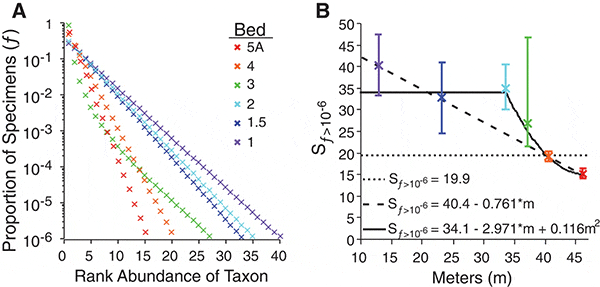
Полученные данные позволили отвергнуть гипотезу о том, что кризис развивался постепенно в течение всего рэтского века (эта гипотеза соответствует наклонной пунктирной линии на рис. 3B). Не подтвердилась и другая гипотеза, согласно которой до самого «рокового» рубежа триаса и юры всё было спокойно, и массовое вымирание свалилось на биосферу как снег на голову. Как выяснилось, кризисные события начали развиваться внезапно и довольно стремительно, но не на самом рубеже, а за полмиллиона лет до него.
Ученые пришли к выводу, что постепенное потепление, связанное с ростом концентрации углекислого газа, могло быть важной, но не единственной причиной кризиса. Возможно, произошло еще какое-то более внезапное событие, ускорившее наступление кризиса (есть данные, согласно которым на землю примерно в это время упал астероид — размером несколько поменьше того, что прикончил динозавров 65 млн лет назад). Таким событием могла быть и резкая активизация вулканической активности на планете. То, что это событие имело глобальный масштаб, следует из того, что позднетриасовый биотический кризис регистрируется в разных частях планеты, и не только для наземной биоты, но и для морских водорослей и мелководных фаун.
Однако можно предположить, что никаких внезапных событий всё-таки не было, а всё дело в «пороговой» реакции биоты на постепенное потепление. В какой-то момент температура поднялась настолько, что оказалась выше некоторого порогового значения, к которому могли приспособиться тогдашние экосистемы. Чтобы проверить эти гипотезы, необходимы дополнительные данные.
Из «дополнительных материалов» к статье можно узнать, что авторы продолжили свой анализ и выше по разрезу, то есть проанализировали флору следующего, шестого слоя, который, скорее всего, относится уже к началу юры, к геттангскому веку (Hettangian). На рис. 4 показана резкая смена флоры, произошедшая при переходе от пятого к шестому слою, то есть от триаса к юре.
|
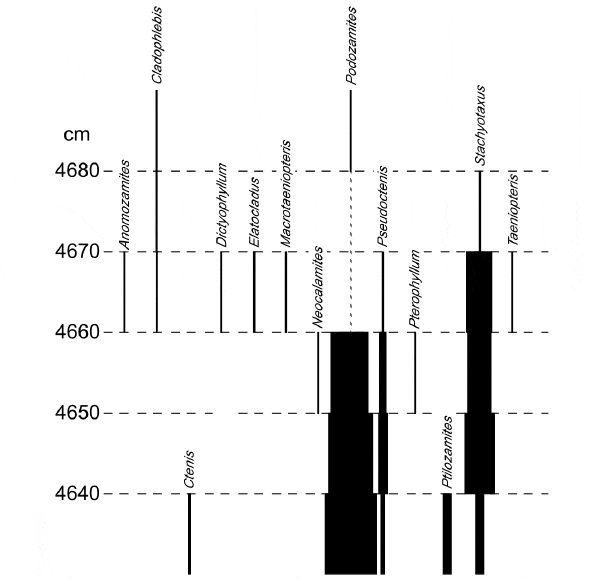
Растительное сообщество, сменившее в начале юры старую, триасовую флору, отличалось гораздо более равномерным распределением родовых обилий, то есть было более сложным и разнообразным (рис. 5).
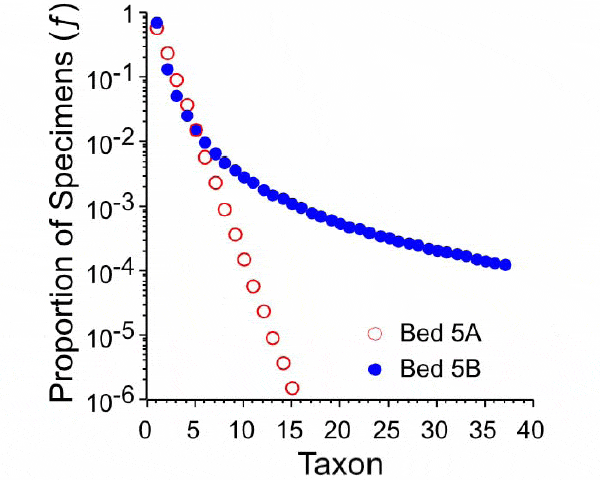
Рис. 5. Математические модели распределения родовых обилий для пятого и шестого слоев (показаны соответственно красными и синими кружками). Модель для пятого слоя (5A), естественно, такая же, как на рис. 3: она показывает очень низкое число «часто встречающихся» родов (около 15). Модель для шестого слоя принципиально иная, она показывает гораздо более высокий уровень разнообразия и гораздо более равномерное распределение родовых обилий, что характерно для богатых, сложных и процветающих сообществ. Рис. из дополнительных материалов к обсуждаемой статье в Science

")