Их открытие, опубликованное в Molecular Cell, объясняет, как единичное событие способно направить клетку в сторону развития рака. «Точное удаление нескольких ограждений ведет к глобальным изменениям в целом геноме, что в конечном итоге приводит к раку», говорит Беверли Эмерсон (Beverly Emerson), профессор лаборатории регуляторной биологии и руководитель проекта.
В норме, сложная сеть ускорителей (факторов роста) и тормозов (опухолевых супрессоров) поддерживает баланс клеточной пролиферации. Опухоли появляются в результате изменений генома, которые активируют гены, вызывающие рак, или при инактивации генов опухолевых супрессоров, что сдвигает этот тонкий баланс в сторону неконтролируемого клеточного роста.
«Очень долго люди пытались понять, каким образом создается молчание генов опухолевых супрессоров в раковых клетках», говорит Михаэль Вичер (Michael Witcher). «Мы выяснили одно из ключевых событий, приводящих к их инактивации, и теперь, вероятно, сможем использовать этот механизм для создания новых методов лечения».
Если вытянуть ДНК одной человеческой клетки, то получится тонкая нитка длиной примерно 2 метра. Чтобы сложить такую длинную молекулу внутри клеточного ядра и содержать ее аккуратно организованно, на ДНК нанизываются белки гистоны, и она сворачивается в высоко конденсированную структуру, называемую гетерохроматином. В областях генной активности хроматин немного разворачивается, чтобы сделать ДНК доступной для регуляторных белков.
В самых различных видах рака, включая рак груди, легких, печени, поджелудочной, множественную миелому и лимфому, ген опухолевого супрессора р16 скрыт глубоко внутри гетерохроматина. В результате с него не идет транскрипция, и р16 не может следить за клеточным ростом.
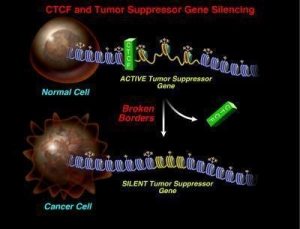
Исследователи уже давно знали, что иногда р16 подавляется задолго до того, как клетка становится раковой. Однако почему именно этот участок помечается химическими маркерами, что приводит к тугому закручиванию и делает эту область недоступной, оставалось загадкой.
Ответ в основном искали в непосредственном окружении данного гена, но всегда оставались с пустыми руками. Когда Вичер расширил свое поле зрения чуть выше данной области, он обнаружил сайт связывания для CTCF, короткого CCCTC-связывающего фактора, который формирует центральный элемент молекулярной конструкции, которая ограждает гетерохроматин от остального генома. «Мы обнаружили нарушение связывания данного белка с несколькими сайтами связывания во многих типах рака, что приводит к коллапсу молекулярного разграничения», говорит он. «Если пропадает граница, то прилежащий гетерохроматин захватывает соседние гены и подавляет их».
Дальнейшие эксперименты показали, что CTCF пропадает из-за недостатка химической модификации, называемой на лабораторном жаргоне "PARlation", что означает поли-АДФ-рибозилирование (poly(ADP-ribosyl)ation), которое позволяет белку связываться с определенным сайтом генома. «Без этой модификации, CTCF не способен формировать комплекс, необходимый для регуляции р16 и другого опухолевого супрессора - RASSF1A. Это объясняет, почему при раке груди в клетках подавлены гены как р16, так и RASSF1A», говорит Вичер.
«Мы считаем, что дестабилизация специфических хромосомных границ и потеря молекулярного ограждения при искажении функций CTCF могут быть главным механизмом инактивации генов опухолевых супрессоров, что инициирует опухолеобразование различных форм рака человека», говорит Эмерсон.

")