|
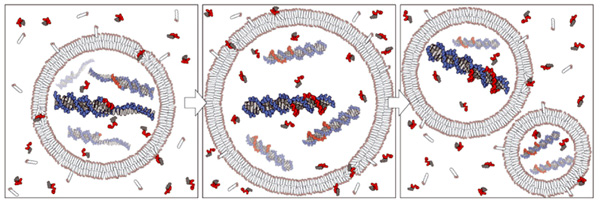
Американские биологи сделали важный шаг к пониманию начальных этапов зарождения жизни. Им удалось создать «протоклетку» с оболочкой из простых липидов и жирных кислот, способную втягивать из окружающей среды активированные нуклеотиды — «кирпичики», необходимые для синтеза ДНК. Протоклетка не может самостоятельно осуществлять матричный синтез (репликацию) ДНК от начала и до конца, но успешно справляется с важнейшими этапами этого процесса, причем все реакции идут без участия каких-либо белков или других сложных биологических молекул-катализаторов.
Другие эксперты считают более вероятным, что первые организмы были автотрофами, то есть не нуждались в готовой органике и синтезировали ее сами из углекислого газа и других простых веществ, используя для этого энергию окислительно-восстановительных реакций (хемоавтотрофы) или света (фотоавтотрофы). Впрочем, идея о первичности фотоавтотрофов представляется сомнительной, поскольку данные сравнительной геномики убедительно свидетельствуют о более позднем появлении фотосинтеза по сравнению с некоторыми типами хемоавтотрофного метаболизма, такими как метаногенез и анаэробное окисление метана.
Молекулярные данные, однако, пока не дают внятного ответа на вопрос о том, кто появился раньше — гетеротрофы или хемоавтотрофы. В пользу первичности гетеротрофов свидетельствует, прежде всего, тот очевидный факт, что их обмен веществ в целом устроен проще. Использовать готовую органику для построения собственных клеток должны уметь все живые организмы, но автотрофам нужно вдобавок эту органику самим синтезировать из простых молекул. Логично предположить, что способность к связыванию CO2 и синтезу органики развилась позже, как «надстройка» над гетеротрофным метаболизмом.
Однако выдвигаются и серьезные доводы против идеи о первичности гетеротрофов. Один из них состоит в том, что поскольку все живые организмы размножаются в геометрической прогрессии, то самая первая появившаяся на планете гетеротрофная форма жизни съела бы весь первичный бульон, сколько бы его ни было, за ничтожный по геологическим меркам срок. Она бы просто не успела за это время пройти весь путь эволюционного развития, необходимый для превращения гетеротрофного организма в автотрофный. На это можно возразить, что «бульон» понемногу подпитывался органикой, образующейся, например, в ходе геохимических процессов в недрах планеты.
Другой аргумент отвести труднее. Мембраны (оболочки) современных клеток состоят из фосфолипидов, и эти мембраны практически непроницаемы для полярных и заряженных молекул, в том числе для сложных органических соединений, таких как сахара или нуклеотиды. Чтобы транспортировать эти молекулы через мембрану, у современных клеток имеется набор специальных транспортных белков. На заре жизни таких белков, конечно, не могло быть. Следовательно, протоклетка просто не могла получать сложную органику из внешней среды. Она должна была довольствоваться теми простыми неорганическими молекулами, которые способны проходить через фосфолипидную мембрану без посторонней помощи. Вывод: первые живые клетки были автотрофами.
Статья американских биологов, опубликованная 4 июня на сайте журнала Nature, представляет собой весьма успешную попытку отвести этот аргумент противников «гетеротрофной теории». Авторы исходили из того, что мембрана протоклетки вовсе не обязательно должна была состоять из тех же самых липидов, что и мембраны современных клеток. Кстати, первичным «веществом наследственности» тоже вовсе не обязательно должны были быть ДНК или РНК в их нынешней форме. Устойчивые двухслойные мембраны (и пузырьки, окруженные такими мембранами) получаются из множества различных липидов, жирных кислот, спиртов и других амфифильных соединений (то есть имеющих полярную гидрофильную «голову» и гидрофобный углеводородный «хвост»). Такие молекулы в воде сами собой могут собираться в двухслойные пленки-мембраны: гидрофобные хвосты поворачиваются внутрь, подальше от воды, а гидрофильные «головы» торчат наружу, образуя оба поверхностных слоя мембраны.
Фосфолипиды — молекулы довольно сложные. Мембраны протоклеток, скорее, должны были собираться из более простых амфифильных соединений, которые могли образовываться абиогенным путем.
Авторы изучили свойства маленьких пузырьков (размером в сотни нанометров, что сравнимо с самыми мелкими живыми клетками), окруженных мембранами из различных жирных кислот. Вначале они пытались выяснить, от чего зависит проницаемость мембран для простых органических соединений, таких как сахар рибоза (этот сахар — одна из необходимых составных частей нуклеотидов, из которых, в свою очередь, собираются молекулы РНК и ДНК). Выяснилось, что мембраны, сделанные из простых жирных кислот, пропускают рибозу немного лучше, чем фосфолипидные мембраны, но все-таки плохо.
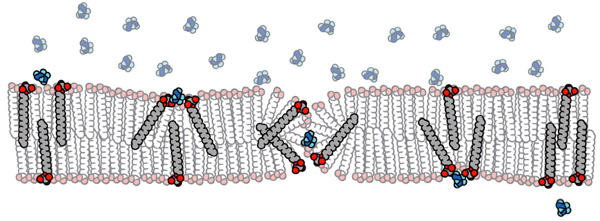
Однако проницаемость резко возрастает, если использовать смесь жирной кислоты с моноэфиром этой же кислоты и глицерина. Многочисленные эксперименты показали, что проницаемость мембраны зависит прежде всего от формы молекул, из которых она сделана: чем больше «голова» молекулы по отношению к длине «хвоста», тем выше проницаемость. Например, у жирных кислот роль «головы» играет карбоксильная группа (–COOH), маленькая по размеру. Длинные гидрофобные «хвосты» в толще мембраны располагаются тесно и плотно слипаются друг с другом. У глицеринового эфира той же жирной кислоты роль «головы» играет молекула глицерина, гораздо более крупная. Из-за этого гидрофобные «хвосты» в толще мембраны размещаются более свободно, и вся конструкция в целом оказывается более рыхлой, текучей и подвижной. На основе проделанных экспериментов авторы предложили теоретическую модель прохождения заряженных молекул через мембраны (см. рисунок).
|
Авторы нашли несколько вариантов состава мембраны, при которых ее проницаемость для рибозы оказывается высокой. Дальнейшие эксперименты проводились с двумя из этих вариантов. Первый из них — смесь миристолеиновой кислоты (Myristoleic acid) с ее же глицериновым моноэфиром (glycerol monoester of myristoleic acid). Эта смесь дает устойчивые пузырьки с хорошей проницаемостью, но у нее есть один недостаток: миристолеиновая кислота содержит 14 атомов углерода и одну двойную связь, и ее присутствие в «первичном бульоне» в достаточно высоких концентрациях считается маловероятным. Второй вариант — смесь декановой кислоты (Decanoic acid) с соответствующим глицериновым моноэфиром и декановым спиртом. Эта смесь ближе к реальности (то есть к тому, что могло быть в первичном бульоне), потому что в декановой кислоте всего 10 атомов углерода и нет двойных связей.
Затем авторы приступили к изучению проницаемости этих пузырьков по отношению к активированным нуклеотидам — тем «кирпичикам», из которых клетка собирает молекулы РНК и ДНК. Если реальные протоклетки были гетеротрофами, такие нуклеотиды должны были составлять их главную «пищу». Современные клетки используют нуклеотиды с тремя присоединенными к ним остатками фосфорной кислоты (нуклеотид-трифосфаты). Однако нуклеотид-трифосфаты, как выяснилось, наотрез отказываются проходить сквозь любые липидные мембраны. Причина в том, что они несут слишком сильный отрицательный заряд. У нуклеотид-дифосфатов и нуклеотид-монофосфатов заряд меньше, и им удается пройти сквозь миристолеиновые и декановые мембраны, но из таких «кирпичиков» ДНК сама собой не синтезируется.
Однако и здесь нашелся обходной путь. Нуклеотиды можно активировать иным способом — присоединив к ним вместо трех фосфатов один фосфат и молекулу имидазола (имидазол — простое органическое соединение, широко распространенное в живой природе и представляющее собой кольцо из трех атомов углерода и двух атомов азота; имидазол является составной частью одной из 20 «канонических» аминокислот — гистидина). Нуклеотиды, активированные имидазолом, годятся для синтеза ДНК и РНК, но имеют только один отрицательный заряд, а не четыре, как нуклеотид-трифосфаты. Такие нуклеотиды уже применялись ранее в экспериментах по синтезу нуклеиновых кислот без участия ферментов.
Многие исследователи допускают, что на заре жизни для синтеза нуклеиновых кислот могли использоваться не нуклеотид-трифосфаты, как теперь, а нуклеотиды, активированные имидазолом. Такие нуклеотиды даже лучше справляются с данной работой, чем нуклеотид-трифосфаты, особенно при отсутствии белков-катализаторов. Авторы обсуждаемой статьи добавляют к этому еще одно соображение: они предполагают, что переход от нуклеотидов, активированных имидазолом, к менее эффективным нуклеотид-трифосфатам был обусловлен необходимостью предотвратить утечку нуклеотидов из клетки (нуклеотид-трифосфаты, как мы помним, сквозь мембраны не проходят). Это, конечно, произошло уже тогда, когда клетки научились сами синтезировать строительные блоки для синтеза нуклеиновых кислот и перестали «всасывать» их извне.
Как и ожидалось, нуклеотиды, активированные имидазолом, достаточно свободно проходили сквозь миристолеиновые и декановые мембраны. Этот успех вдохновил авторов на попытку создания искусственной протоклетки, которая «питалась» бы активированными нуклеотидами и осуществляла матричный синтез (репликацию, копирование, размножение) молекул ДНК или РНК без помощи ферментов.
На сегодняшний день химики уже добились кое-каких успехов в изучении неферментативной репликации нуклеиновых кислот. Однако условия, необходимые для прохождения полного цикла репликации без помощи белков, пока еще не удалось подобрать. Остались две главные нерешенные проблемы. Во-первых, пока не найдены условия, в которых шел бы сам собой матричный синтез любой молекулы ДНК или РНК независимо от последовательности нуклеотидов в матрице. Одни последовательности удается реплицировать, другие нет. Во-вторых, чтобы процесс самопроизвольной репликации начался, нужна «затравка» — праймер. Это значит, что если взять простую одноцепочечную молекулу ДНК или РНК, то на такой матрице без помощи ферментов репликация не начинается. Начинать ее приходится все-таки с использованием ферментов. Но если часть нуклеотидов второй (комплементарной) цепочки уже стоит на своих местах, то процесс репликации может в определенных условиях продолжаться без помощи ферментов. И это уже немало.
Если бы полный цикл неферментативной репликации НК был уже открыт, то авторы обсуждаемой статьи, видимо, подошли бы вплотную к созданию настоящего живого организма. Ну а пока им пришлось довольствоваться тем, что есть. Они взяли короткие молекулы ДНК с затравкой и с недореплицированным «хвостиком», состоящим из 15 нуклеотидов Ц (цитидинов). Молекулы были помещены внутрь мембранных пузырьков.
Эти пузырьки с начинкой — модельные протоклетки — поместили в среду, оптимальную для неферментативного синтеза ДНК (pH 8,5, температура 4°C, плюс еще два простых органических соединения, теоретически совместимых с представлениями о первичном бульоне). После этого протоклетки стали получать «пищу» — активированные нуклеотиды. Официальное название «корма»: 2'-амино-2',3'-дидеоксигуанозин-5'-фосфоримидазол. Время от времени часть протоклеток извлекалась из раствора, чтобы посмотреть, как идет репликация.
Шла она хорошо, хоть и медленно. В конце концов все протоклетки справились с задачей, то есть закончили репликацию недореплицированных молекул ДНК, пристроив к каждому из 15 цитидинов (Ц) комплементарный ему гуанозин (Г). На это у них ушло 24 часа, по 96 минут на нуклеотид. В настоящих живых клетках репликация ДНК осуществляется в десятки миллионов раз быстрее, но ведь там есть сверхэффективные катализаторы — ферменты.
Полученные результаты показывают, что первые живые клетки все-таки могли быть гетеротрофами. А еще они показывают, что уже в самом ближайшем будущем ученые, по-видимому, смогут воспроизвести в лаборатории все ключевые этапы зарождения жизни из неживой материи.

")