До сих пор считалось, что принадлежность термита к той или иной касте зависит только «от воспитания», то есть от того, чем его кормят другие термиты и какие феромоны они при этом выделяют. Японские и австралийские исследователи показали, что так обстоит дело, по-видимому, только у примитивных видов. У более продвинутых форм, таких как распространенный в Японии Reticulitermes speratus, кастовая принадлежность предопределена генетически. Один-единственный ген, расположенный в Х-хромосоме, определяет, по какому из двух возможных путей пойдет развитие насекомого.
У общественных насекомых из отряда перепончатокрылых (муравьев, ос, пчел, шмелей) роль самцов в общественной жизни сведена к минимуму — они нужны только для размножения и больше ни для чего. Все работы по постройке и защите гнезда, уходу за молодью и добыче пропитания возложены на самок. Кем станет данная самка, рабочей особью, отказавшейся от участия в размножении ради заботы о сестрах, или царицей — основательницей новой колонии, зависит исключительно «от воспитания». Второстепенная роль самцов у перепончатокрылых тесно связана с тем обстоятельством, что они развиваются из неоплодотворенных яиц (то есть партеногенетически) и поэтому являются гаплоидными (имеют одинарный набор хромосом). Это делает их, упрощенно говоря, генетически неполноценными существами, на которых лучше не полагаться в сложных и ответственных делах.
У термитов всё по-другому. Самцы у них диплоидные, развиваются из оплодотворенных яиц и наравне с самками служат как рабочими, так и солдатами. У примитивных видов термитов кастовая принадлежность насекомого зависит, как и у муравьев, только от воспитания, и вплоть до весьма поздних стадий индивидуального развития сохраняется возможность выбора. У более эволюционно продвинутых видов, таких как Reticulitermes speratus, судьба насекомого становится предопределенной уже ко времени второй личиночной линьки (см. рисунок).
|
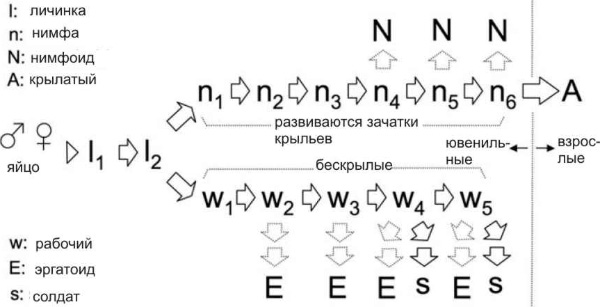
С этого момента развитие насекомого идет по одному из двух путей — «пути нимфы» или «пути рабочего», причем перейти с одного пути на другой уже нельзя.
У нимф образуются зачатки крыльев, и в конце концов они становятся крылатыми взрослыми термитами, способными улететь из гнезда и основать новую колонию.
У рабочих зачатки крыльев не образуются, крылатыми они стать не могут, но некоторые из них после пятой или шестой линьки могут быть призваны на военную службу — стать солдатами.
Термитники живут десятилетиями и, как всякое большое и серьезное предприятие, они располагают средствами для поддержания своей стабильности и жизнеспособности. Гибель царя или царицы в термитнике — дело довольно обычное, и это вовсе не приводит автоматически к смерти всей колонии. В отсутствие размножающихся особей роль «и.о. царя и царицы» могут взять на себя как рабочие бескрылые самцы и самки, так и нимфы с зачатками крыльев. Они приступают к размножению, сохраняя внешний вид ювенильных (неполовозрелых) особей. Размножающихся рабочих называют «эргатоидами», размножающихся нимф — «нимфоидами».
Кроме того, самки всех трех сортов (царицы, нимфоиды и эргатоиды) могут в отсутствие самцов размножаться партеногенетически. Всё потомство при этом оказывается женского пола (у большинства термитов, как и у многих других животных и у людей, самки имеют набор половых хромосом XX, самцы – XY). В отличие от большинства перепончатокрылых, у которых партеногенетическое потомство гаплоидное, у термитов оно диплоидное, то есть имеет двойной набор хромосом. Это происходит за счет простого удвоения гаплоидного хромосомного набора яйцеклетки, поэтому такие термиты оказываются гомозиготными по всем генам (каждый ген представлен двумя идентичными копиями).
Японские и австралийские энтомологи провели серию экспериментов, целью которых было выяснить, чем определяется выбор одного из двух путей развития — воспитанием или генотипом. Для этого они скрещивали между собой в разных комбинациях нимфоидов с эргатоидами, а также заставляли самок размножаться партеногенетически.
Каждой размножающейся паре и каждой партеногенетической самке давали «в помощь» 50 рабочих-самок из другой, неродственной колонии (присутствие рабочих стимулирует откладку яиц). Все отложенные яйца ежедневно забирались из искусственных термитников и отдавались на «воспитание» пятидесяти рабочим-самцам, тоже из неродственной колонии. Таким образом, все личинки воспитывались в одинаковых условиях и в отсутствие размножающихся особей (превращение рабочих в эргатоидов — дело не быстрое, оно занимает 30–40 дней). После третьей линьки ученые регистрировали пол каждой личинки и ее кастовую принадлежность (рабочая или нимфа).
|
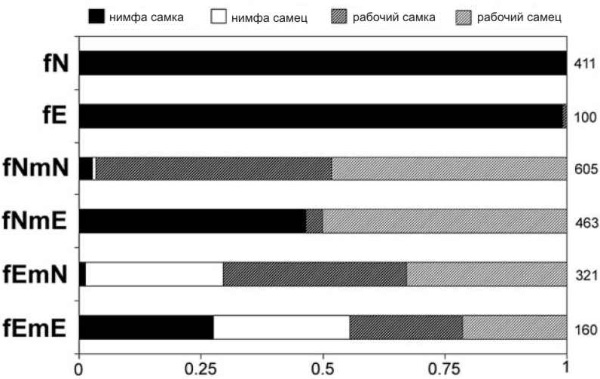
Полученные результаты (см. рисунок) четко показали, что каста зависит не столько от воспитания, сколько от генотипа, причем ключевую роль в определении кастовой принадлежности играет один-единственный ген, локализованный в X-хромосоме.
Результаты, действительно, получились крайне интересные. Несколько упрощая картину, их можно описать следующим образом:
1) партеногенетическое потомство самок-нимфоидов на 100% состояло из самок-нимф;
2) партеногенетическое потомство самок-эргатоидов наполовину погибло, а почти все выжившие оказались самками-нимфами;
3) потомство от скрещивания самок-нимфоидов с самцами-нимфоидами наполовину состояло из самок-рабочих, наполовину — из самцов-рабочих;
4) потомство от скрещивания самок-нимфоидов с самцами-эргатоидами наполовину состояло из самок-нимф, наполовину — из самцов-рабочих;
5) потомство от скрещивания самок-эргатоидов с самцами-нимфоидами на четверть погибло, а остальные 3/4 состояли в равной пропорции из самцов-нимф, рабочих-самок и рабочих-самцов;
6) потомство от скрещивания самок-эргатоидов с самцами-эргатоидами состояло из самок-нимф, самцов-нимф, самок-рабочих и самцов-рабочих в равной пропорции (1:1:1:1).
Простейшим объяснением полученных результатов является следующая генетическая модель. В X-хромосоме имеется ген (исследователи назвали его worker), который существует в двух аллельных вариантах: A и B. Это дает три возможных генотипа для самок и два для самцов:
XAXA — самка-нимфа,
XAXB — самка-рабочая,
XBXB — леталь (погибает),
XBY — самец-нимфа,
XAY — самец-рабочий.
Дополнительные эксперименты показали, что генетическая детерминация кастовой принадлежности не является абсолютно строгой (воспитание может, например, сделать часть генетических самок-нимф самками-рабочими), но в целом каста зависит от генов намного сильнее, чем от воспитания.
Исследователи полагают, что система генетической детерминации каст возникла как надстройка над исходной «воспитательной» системой. Это дало термитам ряд преимуществ. Образно говоря, строгие законы генетики регулируют кастовый состав колонии более надежно, чем менее строгие законы общественной жизни. Например, генетическая детерминация каст гарантирует, что в молодой колонии, пока основавшие ее царь (XBY) и царица (XAXA) еще живы и полны сил, будут появляться на свет только рабочие особи (XAXB и XAY). Это дает возможность молодой колонии расти быстрее, не тратя энергию на производство крылатых особей. Нимфы начнут появляться только после того, как родители-основатели зачахнут или погибнут, и часть рабочих превратится в эргатоидов.
Авторы предложили также эволюционную модель, объясняющую, в какой последовательности должны были появляться и распространяться аллели гена worker. Сначала должен был распространиться аллель A, что привело к появлению генетически детерминированных рабочих-самцов. Поскольку рабочих-самцов стало больше, это автоматически привело к тому, что среди крылатых выросла доля самок. Это, в свою очередь, создало предпосылки для распространения аллеля B, который сделал генетически детерминированными также и рабочих-самок, а заодно и «выровнял» соотношение полов среди крылатых. То, что в результате иногда стали откладываться нежизнеспособные яйца с генотипом XBXB, было своеобразной «платой» за полученные преимущества.
Модель предсказывает, что в процессе перехода от «воспитательной» детерминации каст к «генетической» должны были появляться своеобразные переходные формы термитов с резко неравным соотношением полов в некоторых кастах. Нечто подобное действительно наблюдается у некоторых термитов. В частности, известно несколько видов, у которых все рабочие и (или) все эргатоиды относятся только к одному полу. Вполне возможно, что переход к генетической детерминации кастовой принадлежности за счет появления тех или иных регуляторных аллелей в половых хромосомах происходил (и продолжает происходить) многократно в разных эволюционных линиях термитов.

")