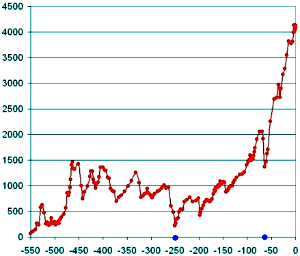
Изменение числа родов морских животных в течение фанерозоя. По горизонтальной оси — время в млн лет назад. Слева — начало фанерозоя, справа — современность. Синими точками отмечены два наиболее катастрофических вымирания — на рубеже палеозоя и мезозоя (250 млн лет назад) и на рубеже мезозоя и кайнозоя (65 млн лет назад). По данным Дж. Сепкоски (1996)
Палеонтологи из США и Австралии на основе статистического анализа 1176 палеонтологических коллекций пришли к выводу, что массовое вымирание на рубеже палеозойской и мезозойской эр привело к радикальному изменению структуры морских сообществ. До этого события в морях преобладали сообщества с простой структурой, слабыми экологическими связями между видами, низким видовым разнообразием и доминированием неподвижных фильтраторов. Великое вымирание уничтожило эти древние сообщества. Когда биота восстановилась после кризиса, новые сообщества оказались более сложными и разнообразными, а экологические связи между видами стали более тесными. В обновленных сообществах стали преобладать подвижные животные.
Палеонтологическая летопись позволяет с большой точностью и детальностью восстанавливать эволюционную историю многих групп животных — особенно тех, у кого есть твердый скелет. Значительно сложнее по палеонтологическим данным судить об эволюции экосистем и биосферы в целом.
Если брать «сырые» палеонтологические данные, то они, казалось бы, однозначно говорят о том, что в течение последних 540 миллионов лет разнообразие животного мира в целом росло, а экосистемы становились сложнее, разнообразнее и устойчивее (последние 540 млн лет — это фанерозойский эон, для которого летопись наиболее полна, так как именно 540 миллионов лет назад появились и быстро размножились животные со скелетом).
Этот рост не был равномерным: он происходил с большим ускорением, а более детальный анализ данных показывает, что ускорение роста происходило скачкообразно. В эволюции фанерозойской морской биоты отчетливо выделяются три переломных момента: 1) великая «радиация» в начале ордовика (460-490 млн лет назад), когда происходило массовое появление классов и отрядов животных; 2) величайшее из всех массовых вымираний, произошедшее на рубеже палеозоя и мезозоя (251 млн лет назад); 3) второе по масштабу массовое вымирание на границе мезозоя и кайнозоя (65 млн лет назад). По нашим данным, после каждого из этих кризисов скачкообразно росла средняя продолжительность существования вновь появляющихся родов морских животных (см. А. В. Марков. О механизмах роста таксономического разнообразия морской биоты в фанерозое // Палеонтологический журнал. 2002. № 2. С. 3-13).
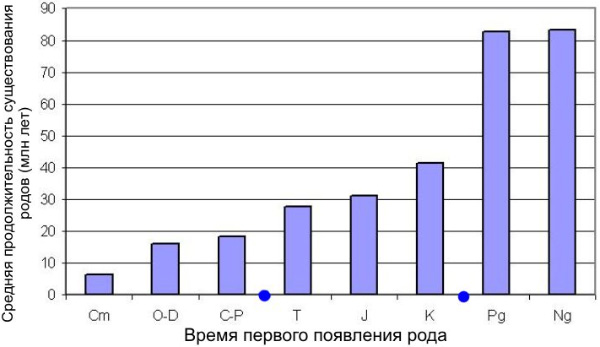
Если сгруппировать роды фанерозойских морских животных по времени их появления, становится заметен ступенчатый рост средней продолжительности их существования. Чем позже появился род, тем он дольше проживет. Синими точками отмечены моменты великих вымираний на рубеже палеозоя/мезозоя и мезозоя/кайнозоя. Обозначения периодов палеозоя: Cm — кембрий, O — ордовик, D — девон, C — карбон, P — пермь; мезозоя: T — триас, J — юра, K — мел; кайнозоя: Pg — палеоген, Ng — неоген. По: Марков, 2002; данные пересчитаны по новой (2004 г.) геохронологической шкале
Суммарное биоразнообразие складывается из трех компонентов: альфа-разнообразие (среднее число видов в одном сообществе), бета-разнообразие (разнообразие типов сообществ в пределах одной биогеографической области), гамма-разнообразие (разнообразие биогеографических областей). Альфа-разнообразие является также простейшей мерой сложности, продвинутости и, если угодно, совершенства экосистем.
Из-за неполноты летописи проследить динамику альфа-разнообразия гораздо труднее, чем динамику суммарного биоразнообразия. По имеющимся оценкам, полученным еще в 70–80-е годы (и с тех пор эти оценки не стали точнее, несмотря на немалые усилия исследователей), среднее число видов в сообществе скачкообразно возрастало дважды: в начале ордовика и на рубеже мезозоя и кайнозоя, то есть на первом и третьем из трех перечисленных критических рубежей.
Менялся также и качественный состав морских сообществ. В начале фанерозоя преобладали неподвижные, прикрепленные организмы — фильтраторы, такие как морские лилии, брахиоподы, мшанки. Хищных, активно плавающих и ползающих, а также роющих форм было сравнительно мало. В дальнейшем от кризиса к кризису ситуация менялась, причем тоже скачкообразно: становилось больше подвижных форм, в том числе роющих, росло относительное разнообразие хищников. Виды вымирали всё реже, а это значит, что сообщества и отдельные виды становились более устойчивыми ко всевозможным переменам среды.
Эти факты, казалось бы, ясно говорят о том, что в течение фанерозоя морские сообщества становились разнообразнее, сложнее, совершеннее и устойчивее. Однако в последние годы многие палеонтологи усомнились в достоверности этой картины. Дело в том, что неполнота летописи, как выяснилось, по целому ряду причин сильно растет с возрастом. Например, древние осадочные породы чаще оказываются литифицированными, то есть превратившимися в сплошной каменный монолит, по сравнению с молодыми породами, сохраняющими рыхлость и сыпучесть. Из литифицированных пород гораздо труднее добывать ископаемые остатки животных, особенно мелких. Не это ли является причиной того, что палеозойские палеонтологические коллекции оказываются беднее и однообразнее мезозойских и кайнозойских? Может быть, и весь наблюдаемый рост биоразнообразия в течение фанерозоя является артефактом, а в действительности нынешний уровень биоразнообразия был достигнут еще в самом начале фанерозоя и с тех пор не менялся?
Кроме литифицированности есть и другие возможные источники систематических ошибок, которые, в принципе, могут создавать иллюзию роста разнообразия в течение фанерозоя. Это избирательное растворение минеральных скелетов определенного типа (арагонитовых) в древних породах, а также то обстоятельство, что молодых осадочных пород в целом сохранилось больше, чем древних. Есть, однако, и противоположные тенденции. Так, наиболее изученные в палеонтологическом отношении регионы (Европа, Северная Америка) в палеозое находились в экваториальной области, а затем сместились в умеренные широты. Разнообразие на экваторе всегда выше, чем в умеренном поясе, поэтому данная тенденция по идее должна не преувеличивать, а преуменьшать наблюдаемый рост разнообразия в течение фанерозоя.
Все эти проблемы встали перед палеонтологами в последние годы в связи с созданием большой международной базы данных по палеонтологическим коллекциям (The Paleobiology Database), и на сегодняшний день ситуация представляется весьма запутанной. Специалисты разделились на два лагеря: одни считают, что наблюдаемой картине доверять вообще нельзя, другие доказывают, что все-таки можно, поскольку выявленные источники ошибок отчасти компенсируют друг друга, а отчасти — не столь существенны, как утверждают оппоненты.
Сам я во втором лагере, и поэтому появление в последнем номере журнала Science статьи с новыми доказательствами «нашей» правоты не могло меня не порадовать.
Питер Вагнер, Мэттью Косник и Скотт Лидгард проанализировали данные по 1176 коллекциям из The Paleobiology Database, для которых в базе данных есть сведения о численности каждого вида. Согласно имеющимся экологическим теориям, о структуре сообщества можно судить по характеру количественного распределения видовых обилий (РВО). Самый простой способ — подсчитать равномерность этого распределения. Для развитых, устойчивых сообществ характерно более равномерное РВО. Это значит, что численности разных видов, входящих в сообщество, не очень сильно отличаются друг от друга. Для примитивных, угнетенных, слаборазвитых сообществ характерно резкое преобладание одного или немногих видов, а все остальные виды присутствуют в ничтожном количестве. Считается, что это более тонкая и надежная мера сложности сообщества, чем простой подсчет видов в нём (то есть измерение альфа-разнообразия).
Ранее уже было показано (M. G. Powell, M. Kowalewski. 2002. Increase in evenness and alpha diversity through the Phanerozoic: Comparison of Early Paleozoic and Cenozoic marine fossil assemblages; полный текст Pdf, 230 Кб // Geology. V. 30: P. 331-334), что в течение фанерозоя равномерность РВО в палеонтологических коллекциях растет. Это, казалось бы, говорит о прогрессивном развитии, совершенствовании структуры сообществ. Но это может быть и артефактом, следствием более плохой сохранности древних ископаемых. К тому же, высокая неравномерность РВО в древних коллекциях может свидетельствовать о том, что низкое альфа-разнообразие в палеозое — тоже артефакт. Дело в том, что если взять небольшую случайную выборку из «неравномерной» коллекции, то в выборку попадет меньше видов, чем в том случае, когда исходная коллекция является «равномерной».
Поэтому очень важно было найти какой-то более надежный критерий сложности сообществ, чем простое альфа-разнообразие (число видов) или равномерность РВО. Авторы показали, что таким критерием может служить соответствие РВО одному из нескольких стандартных математических распределений, каждое из которых имеет свою экологическую интерпретацию.
Согласно имеющимся экологическим теориям, если в примитивном сообществе отношения между видами просты (например, сводятся к одной лишь конкуренции за жизненное пространство, причем новые виды, проникающие в сообщество, не создают новых ниш и не способствуют расширению общего экологического пространства, занимаемого сообществом), то РВО должно соответствовать одной из нескольких простых математических моделей. Например, геометрическому распределению, при котором численности видов убывают в геометрической прогрессии от самого массового вида к самому редкому (допустим, самому массовому виду принадлежит 1/2 всех особей в сообществе, второму по массовости — 1/4, третьему — 1/8 и т. д.).
Если сообщество более совершенно, если между видами существуют сложные взаимоотношения, если новые виды расширяют занимаемое сообществом экологическое пространство и создают новые ниши, тем самым способствуя дальнейшему росту альфа-разнообразия — то в этом случае РВО должно соответствовать одной из более сложных моделей, например лог-нормальному распределению.
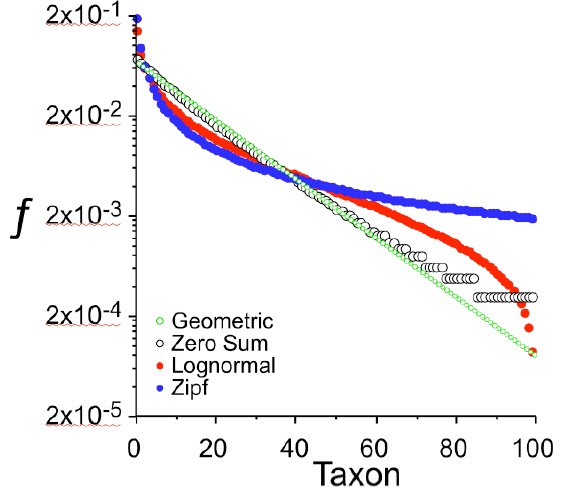
Распределение видовых обилий в «простых» и «сложных» сообществах при одном и том же числе видов (100) и одинаковой равномерности РВО. Это не эмпирические данные, а чистая теория. Простые модели: Geometric и Zero Sum, сложные — Lognormal и Zipf. По горизонтальной оси — порядковые номера таксонов (видов или родов) от первого до сотого, причем таксоны выстроены в порядке убывания их численности. По вертикальной оси — доля таксона в общей численности сообщества. Рис. из дополнительных материалов к статье в Science
Вагнер, Косник и Лидгард показали, что в палеозое наблюдается примерно равное соотношение «простых» и «сложных» сообществ (то есть сообществ, у которых РВО соответствует простым и сложным моделям). Однако после великого вымирания на рубеже палеозоя и мезозоя ситуация резко меняется: с этого момента и вплоть до современности «сложных» сообществ становится в 2-3 раза больше, чем «простых».
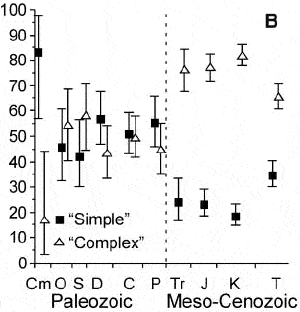
Соотношение «простых» и «сложных» сообществ в течение фанерозоя. Время идет слева направо. Обозначения периодов: Cm — кембрий, O — ордовик, S — силур, D — девон, C — карбон, P — пермь, Tr — триас, J — юра, K — мел, T — третичный (палеоген + неоген). По вертикальной оси — доля коллекций с простыми (квадратики) и сложными (треугольнички) распределениями видовых обилий среди коллекций данного возраста. Вертикальная пунктирная линия — массовое вымирание на рубеже палеозоя и мезозоя. Рис. из статьи в Science
Авторы убедительно показали, что это скачкообразное усложнение сообществ не является артефактом. Например, различия в степени литифицированности молодых и древних пород не могли повлиять на результат, потому что если рассматривать по отдельности коллекции, происходящие из литифицированных и нелитифицированных пород, то в обеих группах коллекций результат получается тот же самый. Аналогичным образом авторы проверили и другие возможные источники ошибок.
Таким образом, после великого вымирания на рубеже палеозоя и мезозоя произошла качественная перемена в структуре морских сообществ. В новых сообществах между видами сложились более сложные взаимоотношения, не сводящиеся к одной лишь конкуренции и включающие элементы сотрудничества. В обновленных морских экосистемах стали преобладать животные, способные расширять экологическое пространство сообщества, создавая ниши для других видов, и способные сами извлекать выгоду из тех изменений среды, которые осуществляются другими членами сообщества.
Это не могло не привести к росту среднего числа видов в сообществе, то есть альфа-разнообразия. Анализ РВО в древних сообществах подтвердил, что наблюдаемый в фанерозое рост альфа-разнообразия, скорее всего, не является артефактом, причем значительный рост этого показателя должен был произойти на рубеже палеозоя и мезозоя, хотя «сырые» данные по числу видов в коллекциях этого не показывают.
Это, в свою очередь, говорит о том, что и наблюдаемый рост общего суммарного биоразнообразия в течение фанерозоя (см. первый рисунок) тоже, скорее всего, не является артефактом.
Таким образом, в течение последних 540 миллионов лет прогрессивное развитие имело место не только на уровне отдельных групп морских животных, но и на уровне целых сообществ, и на уровне всей морской биоты в целом.

")