Существует множество гипотез и теорий, объясняющих происхождение полового размножения, но очень мало экспериментальных данных, подтверждающих или отвергающих эти идеи. Опыты с коловратками класса Monogononta, способными размножаться как бесполым, так и половым путем, подтвердили теорию, согласно которой важным фактором эволюции полового размножения является разнообразие условий среды.
|
|
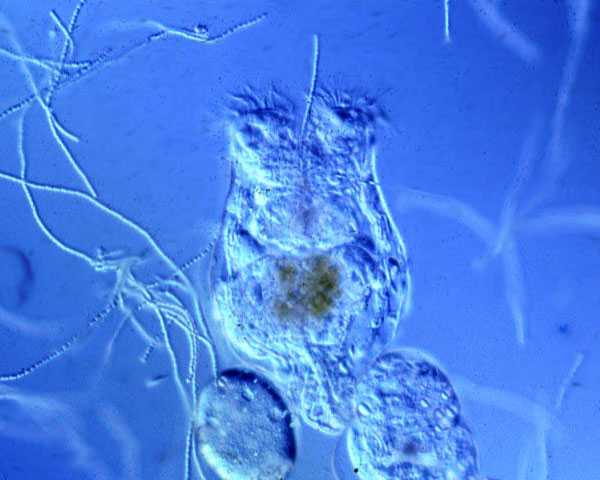
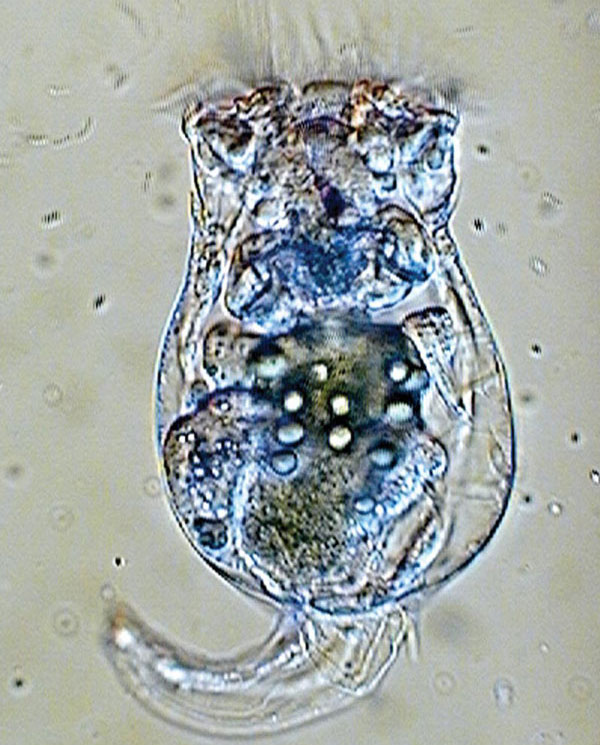
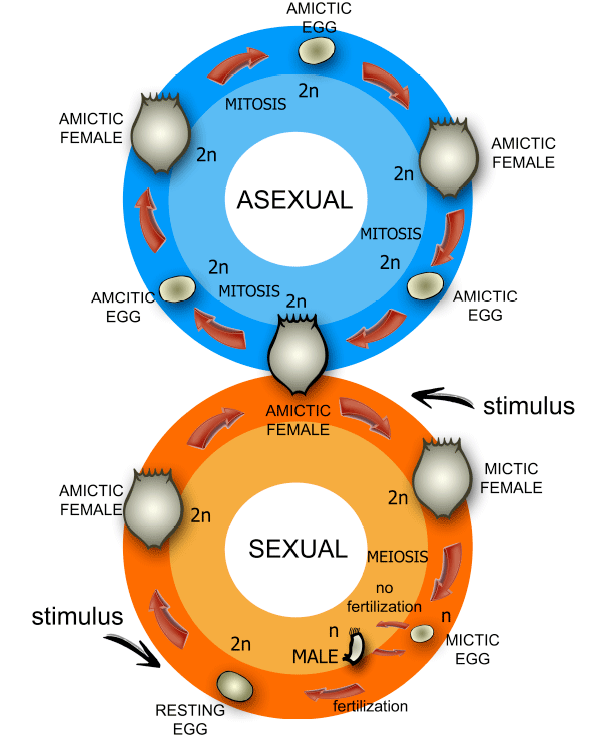
Коловратки класса Monogononta (однояичниковые) способны как к бесполому (партеногенетическому), так и к половому размножению. При низкой плотности популяции в ней, как правило, присутствуют только самки, производящие так называемые амиктические (партеногенетические) диплоидные яйца, из которых без оплодотворения выводится следующее поколение самок. При высокой плотности часть самок начинает производить гаплоидные яйца, из которых выходят маленькие непитающиеся самцы. Они спариваются с самками, в результате чего образуются «покоящиеся» оплодотворенные яйца с плотной оболочкой. Из них снова выводятся только самки. Решение о переходе к половому размножению коловратки принимают на основе «чувства кворума» (см.: «Чувство кворума»: принятие коллективных решений в макро- и микромире, «Элементы», 02.04.2009). Это происходит, когда концентрация веществ, выделяемых самками, превышает определенный порог. Поэтому, чтобы спровоцировать самку на половое размножение, достаточно поместить ее в воду, взятую из аквариума, где плотность популяции коловраток высока.
У коловраток Brachionus обнаружена наследственная изменчивость по склонности к тому или другому способу размножения. Это значит, что выбранный коловраткой в данных условиях способ размножения зависит не только от среды, но и от наследственности: встречаются клоны самок, легко и быстро переходящие к половому размножению, тогда как другие клоны делают это менее охотно.
|
Перечисленные особенности биологии коловраток делают их удобным объектом для экспериментального изучения эволюции пола. Существует много теорий и гипотез, объясняющих, почему половое размножение так широко распространено в природе вопреки высокой «цене», которую за него приходится платить (см.: Опыты на червях доказали, что самцы — вещь полезная, «Элементы», 23.10.2009). Однако экспериментальных работ, подтверждающих или опровергающих эти идеи, проведено пока немного.
Согласно одной из теорий, гетерогенность (разнородность) условий среды может сделать половое размножение более выгодным (Pylkov, Zhivotovsky, Feldman, 1998. Migration versus mutation in the evolution of recombination under multilocus selection; Lenormand, Otto, 2000. The evolution of recombination in a heterogeneous environment; Agrawal, 2009. Spatial heterogeneity and the evolution of sex in diploids). Суть идеи в следующем. Допустим, популяция состоит из двух частей (субпопуляций), живущих в разных условиях, причем между частями существует обмен особями (миграция). В каждой из двух субпопуляций отбор будет благоприятствовать разным комбинациям генов (точнее, генетических вариантов — аллелей). Если особь мигрирует из одной субпопуляции в другую, некоторые ее гены (и комбинации генов) окажутся в новых условиях вредными, снижающими приспособленность. Поэтому всем остальным генам данной особи будет выгодно избавиться от этого груза и найти себе более подходящую «компанию», то есть объединиться с генами аборигенов, лучше приспособленными к местным условиям. Этого очень легко добиться при помощи полового размножения, но совершенно невозможно, если вы размножаетесь партеногенезом. Поэтому, если миграции происходят достаточно регулярно, любая мутация, повышающая склонность особей к половому размножению, имеет шанс распространиться в популяции и вытеснить конкурирующий аллель, снижающий частоту (вероятность) полового размножения.
Биологам из Канады и Германии удалось получить экспериментальное подтверждение этой идеи в ходе экспериментов с коловратками Brachionus calyciflorus. Подопытные коловратки были выведены из яиц, собранных в одной природной популяции. Ранее было показано, что в этой популяции есть наследственная изменчивость по склонности к сексу, а раз есть наследственная изменчивость, то возможна и эволюция под действием отбора.
Авторы вывели из собранных яиц 120 лабораторных популяций, изначально примерно одинаковых по своим свойствам и уровню изменчивости. Часть популяций затем выращивали в однородных, часть — в разнородных условиях в течение 14 недель, что соответствует примерно сотне поколений. Численность каждой популяции поддерживалась на уровне около 10 000 особей.
Разнородные условия были смоделированы следующим образом. Популяцию делили на две части (субпопуляции). Одну часть помещали в «богатую» питательную среду, а другую — в «бедную». Богатство среды определялось количеством одноклеточных водорослей, которыми питаются коловратки, а количество водорослей — концентрацией азота в воде. Время от времени в каждом аквариуме часть воды заменяли на свежую питательную среду, чтобы количество пищи оставалось примерно постоянным. Миграции между субпопуляциями осуществлялись путем еженедельного пересаживания части коловраток и их яиц из одного аквариума в другой. Использовали два разных уровня миграции: 1% и 10% особей на поколение.
В «однородных» экспериментах всё делалось точно так же, за исключением того, что среда в обоих аквариумах была одинаковая: либо богатая, либо бедная.
Авторы следили за изменениями частоты полового размножения и «наследственной склонности» к нему в каждой популяции. Реальную частоту определяли по соотношению амиктических (неоплодотворенных) и покоящихся (оплодотворенных) яиц. Изменения «наследственной склонности» (частот гипотетических аллелей, регулирующих склонность к половому размножению) определяли при помощи индивидуального тестирования отдельных клонов коловраток. Из каждой популяции брали по 84 самки и выводили из них небольшие партеногенетические клоны. Затем в воду, где жили эти клоны, добавляли немного воды из аквариума с высокой плотностью самок (как уже говорилось, это стандартный способ сексуальной стимуляции коловраток) и подсчитывали число клонов, перешедших к половому размножению.
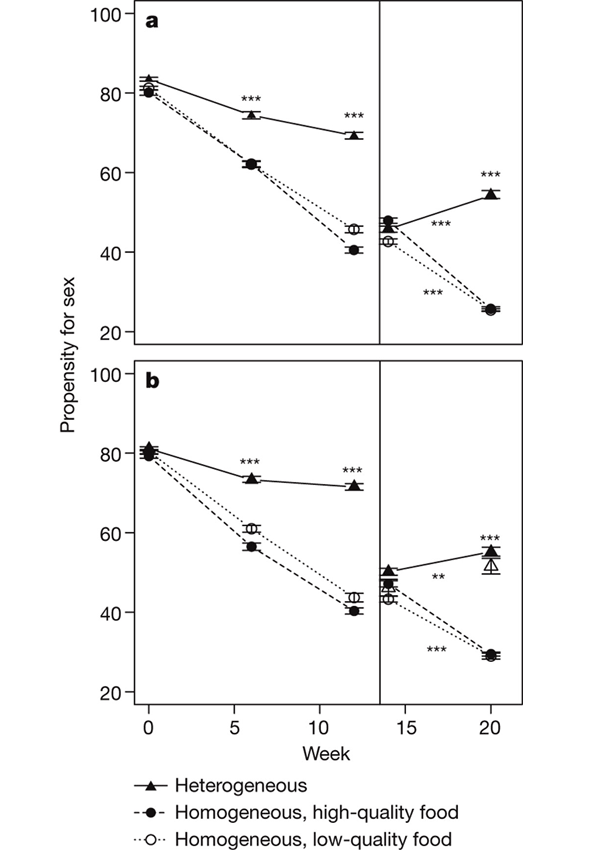
В начале эксперимента подопытные популяции продемонстрировали весьма высокую готовность к сексу: более 80% клонов переходили к половому размножению при соответствующей стимуляции. Через шесть недель в популяциях, живших в однообразной среде, этот показатель упал до 60%, еще через шесть недель — до 40%. Таким образом, в однородной среде коловратки быстро эволюционировали в сторону утраты полового размножения. Этот процесс шел с одинаковой скоростью как в бедной, так и в богатой среде.
В популяциях, живших в разнообразной среде, через 6 и 12 недель после начала эксперимента наблюдался гораздо более высокий уровень полового размножения. Он тоже снизился по сравнению с исходным, но совсем ненамного. По истечении 12 недель не менее 70% клонов в этих популяциях по-прежнему были готовы перейти к половому размножению в ответ на соответствующие стимулы. Уровень миграции (1% или 10% мигрантов на поколение) не повлиял на результаты эксперимента.
Почему частота полового размножения всё-таки снизилась даже в этих популяциях? Возможно, это объясняется тем, что природная популяция коловраток, из которой были выведены лабораторные, живет в еще более разнородных условиях, чем те, что были созданы в эксперименте.
По прошествии 14 недель авторы объединили все экспериментальные популяции, хорошенько перемешали и снова разделили на 120 изолированных линий. Склонность к сексу в этих линиях изначально была около 45–50%. Их опять поместили в однородные или разнородные условия. В первом случае частота полового размножения продолжала снижаться, во втором — начала расти. Таким образом, содержание в разнородных условиях может не только замедлять снижение частоты полового размножения, но и приводить к ее росту.
По-видимому, для таких разнородных условий, которые были созданы в эксперименте, устойчивая или равновесная частота полового размножения (равновесная пропорция клонов, готовых при стимуляции перейти к половому размножению) находится где-то между 55 и 70%. Если частота таких клонов оказывается ниже равновесного уровня, «гены полового размножения» получают селективное преимущество над «генами асексуальности», и наоборот. В однородных условиях равновесный уровень составляет не более 25–30% (до этой отметки упала склонность к сексу у подопытных популяций к концу 20-й недели), а может быть и вовсе равен нулю. Авторы допускают, что если бы они продолжили эксперимент, то, наверное, смогли бы вывести коловраток, полностью утративших интерес к сексу, подобно тому, как это произошло с предками бделлоидных коловраток (класс Digononta или Bdelloidea; см. ссылки внизу).
Авторы считают, что полученные результаты лучше всего объясняются механизмом, описанным выше: половое размножение помогает «разбивать» комбинации генов, подходящие для одних условий, но невыгодные в других. Это косвенно подтверждается, в частности, тем, что коловратки, жившие в течение 15 недель в однородной (бедной или богатой) среде, действительно адаптировались к этим условиям. Адаптированность оценивали по среднему числу потомков, производимых одной самкой в течение жизни. Коловратки, приспособившиеся к богатой среде, производят в ней в среднем около 9 потомков за жизнь, но если их пересадить в бедную среду, их репродуктивный успех падает до 5 потомков. Самки, адаптировавшиеся к бедной среде, производят около 6 потомков, а если их пересадить в богатую среду, их плодовитость падает примерно до 5,5. Следовательно, разные условия действительно способствовали отбору разных аллелей.
|
Одно из альтернативных объяснений состоит в том, что разнородность или переменчивость условий может приводить к общему снижению силы очищающего отбора (то есть отбора, отбраковывающего вредные мутации). Поэтому можно предположить, что в разнородных условиях «гены полового размножения» всё равно остались «вредными», просто отбор стал менее эффективно их отсеивать, и поэтому снижение частоты полового размножения шло медленнее. Но в таком случае скорость снижения этой частоты обязательно должна зависеть от численности популяции. Чем больше популяция, тем слабее в ней эффекты генетического дрейфа (случайных колебаний частот аллелей) и тем эффективнее действие отбора на слабовредные мутации. Чтобы проверить это, авторы провели еще один эксперимент с удесятеренной численностью популяции (100 000 особей вместо 10 000). Это совершенно не повиляло на результаты, а значит, обнаруженный эффект не связан с генетическим дрейфом.
Еще одно возможное объяснение связано с тем, что у коловраток при половом и бесполом размножении образуются разные яйца: в первом случае это яйца с плотной оболочкой, способные переносить неблагоприятные условия, во втором — яйца с тонкой оболочкой, более уязвимые. Может быть, полученные результаты объясняются разной выживаемостью яиц при пересадке из одних условий в другие? Авторы так не считают, потому что в этом случае результаты эксперимента должны были бы сильно зависеть от интенсивности «миграций». Однако такой зависимости выявлено не было.
По-видимому, разнообразие условий среды действительно является важным фактором, не позволяющим большинству организмов отказаться от полового размножения, несмотря на все сложности и затраты, с ним связанные.

")