|

Естественный отбор — главная движущая сила эволюции, поэтому интерес биологов к его изучению вполне понятен. Действие отбора обычно исследуют в лабораторных экспериментах или в ходе многолетних наблюдений в природе.
Первый подход позволяет произвольно менять условия среды, выделяя из бесчисленного множества реальных жизненных обстоятельств отдельные ключевые факторы, от которых зависит направленность отбора. Но условия в эксперименте всегда упрощены и идеализированы. Из-за этого возникает опасность упустить что-то важное и в итоге получить картину, не соответствующую реальности.
Например, в лабораторных экспериментах на рыбках гуппи было показано, что при усилении пресса хищников отбор способствует замедлению старения у жертв. Наблюдения за теми же рыбками в природе выявили противоположную картину: там, где много хищников, гуппи стареют быстрее. По-видимому, несоответствие возникло из-за того, что в лаборатории рыбки меньше страдали от болезней и паразитов, чем в природе. В результате суммарный баланс векторов отбора оказался другим.
Еще один пример: в лабораторных экспериментах по выработке устойчивости к инсектицидам у насекомых обычно возникает полигенная (зависящая от множества генов) устойчивость, тогда как в природе аналогичный эффект бывает связан с изменениями в одном или немногих генах. В данном случае, скорее всего, сказываются различия в численности популяций: природные популяции заведомо больше лабораторных, и поэтому в них чаще появляются очень маловероятные удачные мутации с сильным эффектом, тогда как маленьким лабораторным популяциям приходится эволюционировать за счет накопления множества мутаций со слабым эффектом (об этих и других исследованиях отбора в природе и лаборатории см.: D. N. Reznick, C. K. Ghalambor, 2005. Selection in Nature: Experimental Manipulations of Natural Populations).
Второй подход, основанный на длительных наблюдениях за эволюционирующими популяциями в природе, может дать потрясающие результаты, но он весьма трудоемок и требует незаурядного терпения и мужества. Яркий пример научного героизма — 30-летнее изучение эволюции галапагосских вьюрков, проведенное Питером и Розмари Грант (Peter R. Grant, B. Rosemary Grant, 2002. Unpredictable Evolution in a 30-Year Study of Darwin's Finches). Супруги Грант в деталях проследили, как вслед за изменениями климата и растительности меняется форма клюва и размер тела у двух видов вьюрков. Полученные Грантами результаты сразу попали в разряд классических, но далеко не каждый биолог способен даже ради такого выдающегося достижения полжизни просидеть на Богом забытом тропическом островке вдали от всех благ цивилизации.
Однако дело можно ускорить, если есть возможность прямо в природе манипулировать факторами, влияющими на направленность отбора, а не ждать, пока само собой что-то изменится. Именно по этому пути и пошли Райан Колсбик (Ryan Calsbeek) и Роберт Кокс (Robert Cox) из Дартмутского колледжа, чья статья опубликована недавно на сайте журнала Nature.
Объектом исследования стали ящерицы анолисы. Род Anolis претерпевает бурную адаптивную радиацию и сегодня включает почти 400 видов, распространенных в Северной и Южной Америке и на островах Карибского бассейна. На разных островах эти ящерицы эволюционируют параллельно, давая сходные спектры видов (об аналогичном явлении у птиц см. в заметке Видообразование на разных островах идет параллельными путями, «Элементы», 15.03.2007). Этих ящериц интенсивно изучают уже много лет, что позволило выявить ряд закономерностей их эволюции. Например, известно, что различия в относительной длине конечностей у разных видов анолисов влияют на скорость их лазания по растениям с разной толщиной стеблей; крупные размеры тела характерны для тех видов, которым приходится драться за ресурсы и территорию, и т. д.
Предполагается, что на эволюцию анолисов, обитающих на материке, решающее влияние оказывают хищники, тогда как островные популяции эволюционируют преимущественно под действием внутривидовой конкуренции. Об этом свидетельствуют такие отличительные черты материковых популяций, как низкая плотность, изобилие пищевых ресурсов, раннее половое созревание, слабый половой диморфизм и высокая смертность взрослых ящериц. Эти признаки обычно характерны для популяций, находящихся под сильным прессом хищников.
Целью обсуждаемой работы был сравнительный анализ влияния хищников и конкуренции на направленность отбора у ящериц Anolis sagrei, обитающих на островах Багамского архипелага. В качестве «рабочих полигонов» использовались пять крошечных уединенных островков площадью от 800 до 2300 кв. м, на которых ящерицы не живут, хотя там имеется подходящая для них растительность и пищевые ресурсы. Работа проводилась в течение двух сезонов (в 2008-м и 2009 году).
Авторы поселили на каждом острове от 40 до 80 взрослых самцов и 150 самок Anolis sagrei, пойманных на самом крупном острове архипелага, Великой Эксуме. Характер отбора изучался только на самцах, потому что смертность самцов и самок зависит от разных причин, и к тому же самки параллельно участвовали в другом исследовании и были для этого подвергнуты хирургическому вмешательству. Перед тем как выпустить ящериц на островок, у каждого самца измеряли длину тела и задних конечностей, а также испытывали его выносливость, заставляя бегать в колесе до изнеможения. Все самцы были индивидуально помечены. Выпускали ящериц в мае, а в сентябре собирали выживших. Маленькие размеры островков позволяли собрать практически всех ящериц, оставшихся в живых по прошествии пяти месяцев. Таким образом, исследователи точно знали, кто погиб, а кто уцелел, и могли оценить направленность и силу отбора по каждому из трех признаков (размер тела, относительная длина задних ног, выносливость).
Два из пяти островков использовались дважды, в 2008-м и 2009 году, поэтому общее число экспериментальных полигонов было равно семи. Авторы манипулировали двумя параметрами — плотностью популяции ящериц и наличием хищников. Зная площадь каждого острова и количество выпущенных ящериц, можно рассчитать плотность животных на квадратный метр подходящей для их обитания (то есть покрытой соответствующей растительностью) территории. Плотность экспериментальных популяций варьировала от 0,09 до 0,30 ящериц на кв. м, что примерно соответствует вариабельности этого показателя в природных популяциях. Что касается хищников, то на двух островах их не было вовсе (над островами натянули сетку для защиты от птиц), на трех других доступ для птиц был оставлен открытым, а еще на двух кроме хищных птиц были змеи — авторы выпустили на эти острова по три багамских полоза (Alsophis vudii), которые любят полакомиться анолисами.
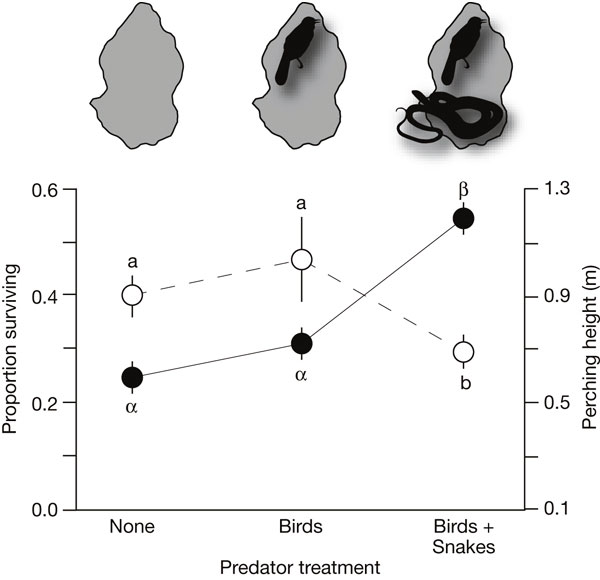
Анализ полученных результатов показал, что присутствие змей (но не птиц) резко повышает смертность ящериц. Кроме того, на тех островах, где были змеи, ящерицы приучились выше забираться по стеблям растений (см. рисунок). Однако никакого влияния хищников на направленность отбора выявлено не было. По-видимому, хищники поедали анолисов неизбирательно, независимо от их размера, длины ног и выносливости. Впрочем, авторы отмечают, что в более долгосрочной перспективе змеи, по-видимому, всё-таки могут влиять на направленность отбора — может быть, не напрямую, а через изменившееся поведение. Например, в присутствии змей могут получить преимущество ящерицы, более ловко карабкающиеся по стеблям, что приведет к выработке дополнительных адаптаций для лазания (см. раздел «Эффект Болдуина» в заметке Гены управляют поведением, а поведение — генами, «Элементы», 12.11.2008).
|
Плотность популяции ящериц, напротив, оказывает весьма сильное влияние на направленность отбора. На островах с высокой плотностью, независимо от наличия хищников, наблюдалось избирательное выживание самых крупных, длинноногих и выносливых самцов.
Таким образом, подтвердилась гипотеза о том, что эффективность естественного отбора у островных ящериц сильнее зависит от плотности популяции (и, следовательно, от конкуренции с себе подобными), чем от пресса хищников. Показательно, что размер тела и относительная длина конечностей — это как раз те признаки, которые быстрее всего меняются в ходе радиации анолисов на островах Карибского бассейна. В дальнейшем было бы логично проверить вторую часть гипотезы, согласно которой отбор в материковых популяциях анолисов направляется уже не плотностью, а хищниками.

")