|
В последние годы коллекция хорошо изученных случаев симпатрического видообразования существенно пополнилась (см. ссылки внизу). Разные пары расходящихся симпатрических видов (и даже целые их «пучки» — species flocks) застигнуты исследователями на разных стадиях процесса расхождения (дивергенции). Наименее изученными остаются самые ранние этапы этого процесса, когда популяция еще остается единой, но внутри нее уже начинает зарождаться частичная репродуктивная изоляция между особями с разными признаками.
Как показало исследование, выполненное сотрудниками Гарвардского, Гавайского и Техасского университетов (США), именно такая ситуация наблюдается в наши дни у бабочек Heliconius cydno alithea, обитающих в западном Эквадоре. Бабочки рода Heliconius защищены от хищников, во-первых, своей ядовитостью, во-вторых, яркой «предупреждающей» окраской. При этом у них широко распространена так называемая мюллеровская мимикрия (Mullerian mimicry) — маскировка одних ядовитых видов под другие (о другом виде мимикрии — бейтсовской, при которой неядовитые виды маскируются под ядовитых, см.: Необычная маскировка у лягушек связана со способностью хищников к обобщениям, «Элементы», 13.03.2006). Мюллеровская мимикрия ускоряет обучение хищников: «обжегшись» на одной из ядовитых бабочек, хищник уже не будет испытывать судьбу, нападая на другие похожие виды. Ранее на бабочках Heliconius было показано (в том числе при помощи экспериментов по переселению бабочек из одних частей ареала в другие), что приспособленность (выживаемость) бабочек-подражателей в природе напрямую зависит от частоты встречаемости бабочек-моделей: из всех подражателей самая высокая приспособленность оказывается у тех, кто маскируется под наиболее массовый вид «моделей» в данной местности. Этот результат, между прочим, является одним из многих экспериментальных подтверждений действенности естественного отбора в природе.
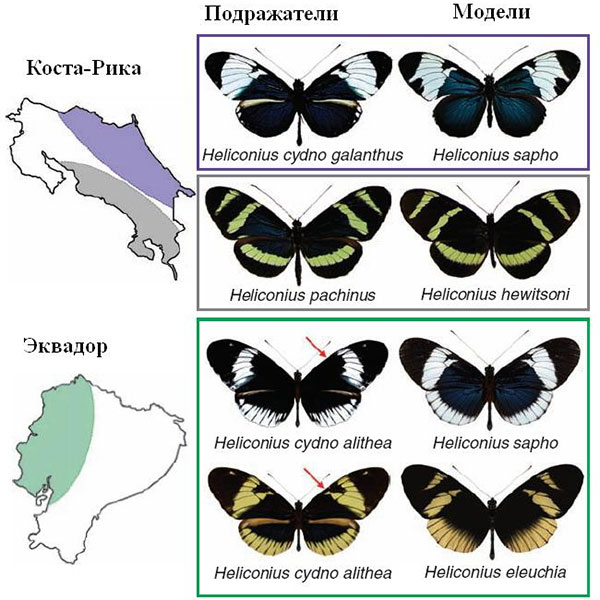
Ранее были подробно изучены две пары подражающих друг другу видов Heliconius cydno galanthus / H. sapho и H. pachinus / H. hewitsoni, обитающие по разные стороны водораздела в Коста-Рике. Первый вид в каждой паре — подражатель, второй — модель; для первой пары характерны белые пятна на крыльях, для второй — желтые. Генетический анализ показал, что виды-подражатели состоят в более близком родстве друг с другом, чем со своими моделями. В лабораторных условиях два вида подражателей способны скрещиваться и давать плодовитое гибридное потомство, хотя при наличии выбора они всегда предпочитают партнеров из своего вида. Межвидовые скрещивания позволили установить, что цвет пятен на крыльях (белый или желтый) определяется двумя аллелями (вариантами) одного-единственного гена, причем белый аллель — доминантный, а желтый — рецессивный. Наследование происходит строго «по Менделю»: при скрещивании белых H. cydno galanthus с желтыми H. pachinus первое поколение гибридов оказывается белым, а во втором наблюдается расщепление 3:1 (три белых бабочки на одну желтую). Природа гена, отвечающего за выбор между желтой и белой окраской, пока не установлена. Известно только его положение на хромосоме, а сам ген фигурирует в литературе под условным названием «локус К».
H. cydno galanthus и H. pachinus — это «хорошо разошедшиеся» виды, в природе не скрещивающиеся и даже не симпатрические: они обитают в разных районах Коста-Рики. Генетический анализ подтвердил, что их генофонды уже довольно давно существуют в изоляции друг от друга.
Иная картина наблюдается в Эквадоре, где тоже есть два вида-модели — белый и желтый, и два подражателя. Однако в данном случае оба подражателя относятся к одному и тому же виду (формально — даже к одному подвиду): H. cydno alithea. Обе модели и вид-подражатель, подразделенный на желтую и белую формы, проживают на одной и той же территории.
|
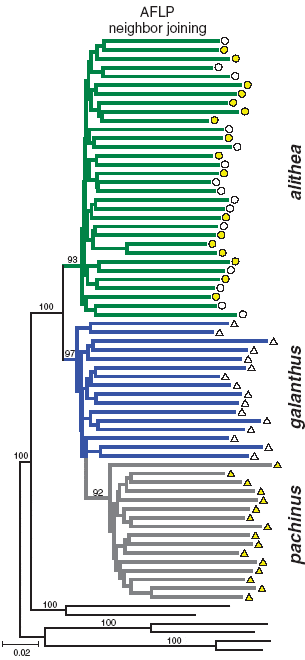
Скрещивая бабочек между собой, авторы убедились, что наследование признака «желтые или белые пятна на крыльях» у H. cydno alithea основано на том же генетическом механизме, что и у видов из Коста-Рики, и отвечает за него тот же самый локус К. Затем авторы экспериментально изучили предпочтения белых и желтых самцов H. cydno alithea при выборе самок. Оказалось, что желтые самцы явно предпочитают желтых самок. Белые самцы, напротив, оказались неизбирательными: они одинаково активно ухаживают и за белыми, и за желтыми самками.
Авторы проанализировали более 800 полиморфных участков генома у белых и желтых бабочек. Полученные результаты свидетельствуют о том, что предпочтение «себе подобных», характерное для желтых самцов, не в состоянии заметно ослабить генетический обмен между формами с разным цветом крыльев. Несмотря на избирательность желтых самцов, желтые и белые бабочки H. cydno alithea фактически образуют единую популяцию. Это проявляется, в частности, в том, что на эволюционных деревьях белые и желтые особи располагаются вперемежку и наотрез отказываются формировать обособленные кластеры.
Таким образом, налицо единая популяция, внутри которой лишь едва наметилось разделение на две репродуктивно изолированные части. Эти части уже дивергировали экологически, то есть разошлись по экологическим нишам (в данном случае — по «подражательным» нишам: они подражают разным видам-моделям). Между ними наметилось и репродуктивное разделение, но оно проявляется пока только в брачных предпочтениях самцов одной из двух форм. Авторы отмечают, что это разделение в будущем вполне может стать (а может и не стать, конечно) основой для полного разделения исходной популяции на два вида. Одних лишь предпочтений желтых самцов для этого мало. Популяция сможет разделиться на два вида только в том случае, если появятся дополнительные факторы, способствующие такому разделению. Авторы упоминают три возможных сценария, которые в будущем могут разделить H. cydno alithea на два вида:
1) Появление еще какого-нибудь признака, кроме цвета крыльев, по которому бабочки будут проявлять положительную избирательность (предпочтение себе подобных) при скрещиваниях. Хорошим кандидатом на такой признак являются вариации в форме и размере черных пятен на передних крыльях (на первом рисунке эти вариации отмечены красными стрелками). Имеющиеся вариации делают бабочек-подражателей более или менее похожими на разные модельные виды, однако их влияние на приспособленность подражателей пока не выявлено. Особи с разной формой черных пятен встречаются с одинаковой частотой среди желтых и белых бабочек.
2) Появление положительной избирательности не только у желтых, но и у белых самцов.
3) Пространственное расхождение ареалов двух видов бабочек-моделей, которое, в свою очередь, может быть вызвано расхождением ареалов их кормовых растений.
Если что-то из перечисленного в будущем произойдет, не исключено, что H. cydno alithea разделится на два вида.
Исследование показало, что репродуктивная изоляция может зародиться в популяции даже в том случае, если никакой явной выгоды эта изоляция никому не дает. Считается, что при экологическом видообразовании (то есть при расхождении будущих видов по разным нишам) важнейшим стимулом для развития изолирующих механизмов является пониженная приспособленность гибридов по сравнению с «чистыми» родительскими формами. Снижение жизнеспособности у гибридов обычно связано со смешиванием родительских адаптивных признаков. Но у бабочек H. cydno alithea при скрещивании желтых особей с белыми никакого смешивания не происходит, потому что адаптивный признак наследуется моногенно, с полным доминированием. Гибридный потомок получает не промежуточный бело-желтый фенотип, который мог бы оказаться неадаптивным, а один из родительских фенотипов в чистом виде — либо белый, либо желтый. Нет никаких оснований предполагать пониженную жизнеспособность потомства от «смешанных» браков по сравнению с потомством от браков «одноцветных».
Почему желтые самцы стали предпочитать желтых самок? Поскольку это предпочтение, очевидно, не дает никаких преимуществ, мы не можем объяснить его развитие действием естественного отбора. Предпочтение сформировалось либо просто случайно, либо автоматически, как побочный продукт каких-то внутренних связей в организме или существовавших ранее биологических механизмов.
Что значит «случайно»? Например, некий гипотетический ген, отвечающий за предпочтение желтых или белых самок, мог случайно оказаться на хромосоме очень близко к локусу К, и поэтому они наследуются сцепленно. Генетические эксперименты подтвердили, что окраска и избирательность действительно наследуются строго сцепленно — однако они не доказали, что эти два признака зависят от разных генов, а не от одного и того же локуса К. К тому же, как бы близко ни располагались друг к другу гены на хромосоме, их сцепленность иногда все-таки должна нарушаться из-за кроссинговера, а это не подтверждается фактами. Поэтому для того, чтобы принять гипотезу «случайной сцепленности», нужно еще ввести дополнительное допущение, согласно которому участок хромосомы, включающий оба гена, не подвергается рекомбинации, то есть кроссинговер внутри него никогда не происходит. Отсутствие рекомбинации может быть вызвано, например, инверсией.
Этот сценарий предполагает случайное совпадение нескольких маловероятных событий. Между тем симпатрическое видообразование — явление весьма распространенное. Поэтому должны быть другие механизмы автоматического зарождения изоляции, не требующие такого редкого стечения обстоятельств.
Не исключено, что локус К может одновременно влиять и на цвет крыльев, и на избирательность самцов. По мнению авторов, это не так уж невероятно — особенно если учесть, что у бабочек одни и те же пигменты могут использоваться и для окрашивания крыльев, и как светофильтры в глазах.
Другая возможность — на мой взгляд, самая очевидная — состоит в том, что самец делает выбор «с оглядкой на себя»: он видит, какого цвета крылья у него самого, и выбирает таких же самок. В этом случае самец-мутант с изменившимся цветом крыльев автоматически сразу начнет предпочитать самок с такой же мутацией. Но тогда остается непонятным, почему выбирают себе подобных только желтые самцы, а белые ухаживают за всеми самками без разбора.
При решении этой загадки следует учитывать, что желтая окраска — рецессивный признак, а белая — доминантный. Поэтому все желтые самцы гомозиготны по локусу К, а большинство белых — гетерозиготны: одна из копий этого гена у них «желтая», другая — «белая». Если предположить, что самцам известен не только цвет крыльев самки, но и ее генотип (гетерозиготна она или гомозиготна по локусу К), то модель «выбора с оглядкой на себя» полностью объясняет наблюдаемую картину. Получается, что и белые, и желтые самцы предпочитают себе подобных, но не по фенотипу (цвету крыльев), а по генотипу: каждый самец выбирает самок, у которых есть такие же, как у него, аллели локуса К (о том, как может осуществляться такое «генетическое тестирование», см. в заметке: Видообразование — личное дело каждого, «Элементы», 15.02.2006).
Так или иначе, окончательного ответа на вопрос, почему желтые самцы предпочитают желтых самок, пока нет. Однако данная работа убедительно показывает, что отдельные «строительные блоки», из которых в дальнейшем может быть построен надежный изолирующий барьер между расходящимися видами, могут формироваться в недрах единой популяции даже в том случае, если никакого адаптивного преимущества подобная изоляция (избирательное скрещивание с себе подобными) пока не дает. Это, конечно, никакое не «заглядывание вперед» и не «движение к заранее намеченной цели» — формированию двух новых видов. Скорее всего, это автоматическая реакция выработавшихся ранее в ходе эволюции механизмов выбора брачного партнера на генетические изменения в популяции (см. также: Стресс способствует близкородственным скрещиваниям, «Элементы», 19.03.2009).
В дополнительных материалах к статье дан список хорошо изученных примеров экологического видообразования. В большинстве этих примеров видообразование происходило в условиях симпатрии. Ниже приводится часть этого списка. Он может оказаться полезным не только для споров с креационистами, утверждающими, что якобы «нет доказанных случаев превращения одного вида в другой», но и для переубеждения тех эволюционистов, которые, оставаясь верными идеям Э. Майра, продолжают считать симпатрический сценарий видообразования крайне маловероятным и «верят» только в аллопатрический сценарий.
Другие изученные примеры симпатрического видообразования, «застигнутого» исследователями на разных стадиях:
1) Рыбы Amphilophus citrinellus и A. zaliosus в озере Апойо в Никарагуа (Для видообразования не нужны барьеры, «Элементы», 13.02.2006).
2) Пальмы Howea forsteriana и H. belmoreana на острове Лорд-Хау в 580 км от восточного побережья Австралии (Эволюция без преград: ботаники нашли новое доказательство видообразования без географических барьеров, «Элементы», 13.02.2006).
3) Мухи Rhagoletis pomonella и наездники Diachasma alloeum, подразделившиеся на «яблоневые» и «боярышниковые» формы (Цепная реакция видообразования, «Элементы», 11.02.2009).
4) Трехиглая колюшка Gasterosteus aculeatus, подразделившаяся на два вида — донный и пелагический — в озерах Канады (Экспериментально подтверждено влияние видообразования на свойства экосистем, «Элементы», 06.04.2009).
5) Вьюрки Nesospiza acunhae и N. wilkinsi на островах Тристан-да-Кунья (Видообразование на разных островах идет параллельными путями, «Элементы», 15.03.2007).
6) Палочник Timema cristinae (Sandoval, 2008).
7) Стеблевой мотылек Ostrinia nubilalis.
8) Рыбы-цихлиды рода Pundamilia (Carleton et al., 2005).
9) Морская улитка Littorina saxatilis (Cruz et al., 2004).
10) Растения Mimulus lewisii и M. cardinalis.
11) Бабочка-листовертка Zeiraphera diniana (Emelianov et al., 2001).
12) Гороховая тля Acyrthosiphon pisum (Frantz et al., 2009).
13) Рыба Gambusia hubbsi (Langerhans et al., 2007).
14) Растение Anthoxanthum odoratum (Snaydon & Davies, 1976).
15) Рыбы Hypoplectrus (Puebla et al., 2007).
16) Рыбы-усачи озера Тана в Эфиопии (Sibbing et al., 1998).

")