|

|

Людям, живущем в средней полосе, трудно представить себе то разнообразие, с которым человек сталкивается, оказавшись в тропическом лесу. На площади в один гектар там нередко произрастает более сотни разных видов деревьев, а на площади в 50 га — более тысячи. Для профессионала-эколога столь высокое видовое разнообразие тропических лесов — настоящая загадка. Ведь, согласно классическим представлениям о принципах организации биологических сообществ, сосуществующие виды должны быть ограничены разными факторами (или иначе говоря — занимать разные ниши). Очевидно, однако, что многочисленные, растущие бок о бок, деревья разных видов на самом деле зависят от одного и того же весьма ограниченного набора ресурсов (прежде всего — света, влаги и элементов минерального питания). Поскольку возможностей разойтись по разным нишам для деревьев тропического леса совсем немного, возникло предположение, что сосуществуют они не вопреки, а благодаря своему экологическому сходству.
Согласно концепции нейтрализма, предложенной недавно Стивеном Хаббелом (Stephen P. Hubbell), разные виды могут длительное время произрастать вместе несмотря на конкуренцию за общие ресурсы, если они экологически идентичны. Под экологической идентичностью Хаббел понимал такое положение, при котором в расчете на одну особь («на душу населения» своей популяции) разные виды имеют равную вероятность выжить, размножиться и заселить новое местообитание (Hubbell, 2001). При этом ни у одного из видов нет устойчивого преимущества в конкурентной борьбе, и в результате все виды сосуществуют.
Вокруг гипотезы нейтрализма непрерывно идут жаркие споры, но поскольку подтвердить или опровергнуть ее прямыми наблюдениями в природе крайне сложно, исследователи вынуждены опираться на косвенные свидетельства. В частности, предполагается, что экологические различия видов можно связать с разными жизненными стратегиями (разными способами приспособления к среде и использования ресурсов). Судить же о разных стратегиях можно по морфологическим особенностям растений (например, по максимальной высоте деревьев данного вида, по размеру их семян, по отношению площади листьев к их массе и т. п.). Если произрастающие по соседству деревья разных видов комбинируются чисто случайно (согласно принципу нейтрализма), то различия между соседями должны быть в пределах случайной выборки из всей совокупности видов данной местности (с учетом, конечно, их разной относительной численности). Если же сосуществованию деревьев разных видов способствует наличие экологических различий между ними (традиционная точка зрения), то соседями по лесу чаще должны оказываться виды с разными жизненными стратегиями.
|

Именно такой подход при изучении сообщества деревьев тропического леса использовали Натан Крафт (Nathan J. B. Kraft) из Отдела интегративной биологии Калифорнийского университета (Беркли, США), Ренато Валенсия (Renato Valencia) из Понтификального католического университета (Кито, Эквадор) и Дэвид Экерли (David D. Ackerly) из Калифорнийского университета. Материалом для их статьи, опубликованной недавно в журнале Science, послужили результаты кропотливых исследований в лесу Ясуни (Yasuni Forest) в восточной части Эквадора. Для данной местности отмечено, видимо, максимальное известное на Земле видовое разнообразие деревьев. Так, на модельном участке леса в 25 га (Yasuni Forest Dynamics Plot) выявлено более 1123 видов деревьев (с диаметром ствола не менее 1 см на высоте груди человека), причем все эти деревья (а их более 150 тыс.) определены до вида, измерены и нанесены на карту.
В качестве характеристик жизненных стратегий использовали такие показатели, как удельная поверхность листьев (площадь листьев, отнесенная к их массе), содержание в листьях азота, средний вес семени, максимальный диаметр на уровне груди (косвенно по этой величине можно судить о максимальной высоте дерева) и плотность древесины. Последний показатель взят из литературы, другие непосредственно оценивались в ходе исследования. Некоторые характеристики получены для всех 1123 видов деревьев (например, диаметр ствола) или почти для всех (удельная площадь листьев), другие — для большей части видов, например содержание азота или размер семян. В качестве мерила пространственной близости деревьев использовалось попадание их в одну учетную площадку 20 ? 20 м.
Нуль-гипотеза (нейтрализм) предполагала случайную комбинацию произрастающих вместе видов. Если же индивидуальные особенности видов имеют значение (традиционные представления о структуре сообществ), то возможны два альтернативных варианта. Во-первых, реально сосуществующие виды могут оказаться экологически более близкими, чем при случайной выборке, в силу того, что абиотические условия среды (например, уровень увлажненности почвы или богатство биогенными элементами) в каждом конкретном месте уже производят некоторый отбор («фильтрацию») видов. Во-вторых, сосуществующие виды, наоборот, могут расходиться по своим характеристикам (жизненным стратегиям) и таким образом снижать интенсивность потенциальной конкуренции. Если справедлива первая альтернатива, экологически сходные виды распределяются в пространстве группами в соответствии с особенностями среды. Если справедлива вторая, то ближайшими соседями чаще оказываются деревья с разными жизненными стратегиями.
|
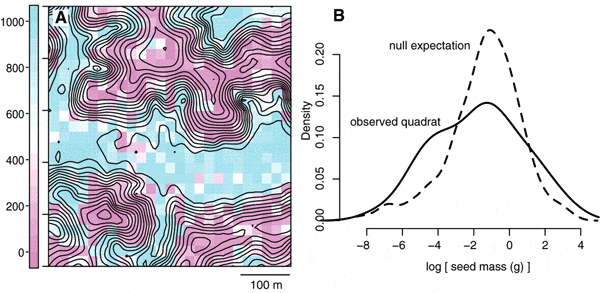
Результаты анализа, проведенного Крафтом и его соавторами, показали, что имеют место оба варианта. На обследованном участке леса есть как группировки в чем-то сходных видов, образованных за счет «фильтрации средой», так и совместное произрастание видов, расходящихся по своим жизненным стратегиям. О «фильтрации средой» свидетельствует хорошая корреляция удельной поверхности листьев с особенностями рельефа — на возвышенных местах преобладают деревья с низкой удельной поверхностью листьев, а в понижениях — с более высокой (см. рис. 1A). О том, что виды, оказавшиеся соседями (попавшие в одну учетную площадку), сильнее расходятся по некоторым признакам, чем того следовало бы ожидать из чисто случайного распределения, свидетельствует обнаруженное «растянутое» распределение признака (см. рис. 1B).
Таким образом, авторы приходят к выводу, что в тропическом лесу жизненные стратегии разных видов деревьев могут демонстрировать как конвергенцию, так и дивергенцию. Гипотеза нейтральности не отвергается, но, возможно, ее недостаточно для объяснения необычайно высокого разнообразия этих сообществ.

")