|
Изучать эволюцию в режиме «реального времени», да к тому же не в лаборатории, а в природе — дело очень трудное. Ведь для этого нужно наблюдать за природными популяциями в течение сотен поколений. Попробуйте-ка получить грант на исследование, если первые результаты ожидаются лишь спустя десятилетия!
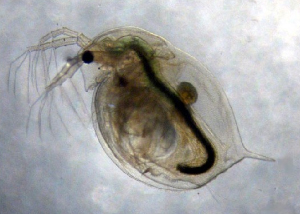
Главной задачей исследователя в этой ситуации становится поиск подходящего природного объекта, позволяющего радикально ускорить получение результата. Для изучения микроэволюционных процессов в системе «паразит—хозяин» замечательным объектом являются пресноводные рачки дафнии и их паразиты — бактерии Pasteuria ramosa. Главное достоинство этой модельной системы состоит в том, что оба ее участника имеют покоящиеся стадии (пропагулы), которые сохраняются в донных отложениях и остаются жизнеспособными в течение многих десятилетий. Извлекая яйца рачков и споры бактерий из последовательных слоев донных осадков, можно восстановить генетическую историю популяций.
Биологи из Бельгии, Швейцарии и Франции опубликовали в последнем номере журнала Nature результаты сравнительного изучения дафний и их паразитов из последовательных слоев донных отложений, образовавшихся в маленьком бельгийском пруду за 30 лет. Толща осадка была условно поделена на 8 слоев толщиной по 2 см. Каждый такой слой образовывался в течение 2–4 лет, что примерно соответствует 10–20 поколениям дафний.
Из каждого слоя были выделены и «оживлены» яйца дафний и споры бактерий. Затем оценивалась инфекционность (заразность) бактерий по отношению к дафниям из разных слоев. Воскрешенные популяции дафний из каждого слоя делили на три группы и пытались заразить бактериями из того же, предыдущего и последующего слоя. Таким образом, оценивались взаимоотношения рачков с «современными», «прошлыми» и «будущими» паразитами.
Оказалось, что наибольшая инфекционность (65% зараженных рачков) характерна для «современных» паразитов, значительно меньшая — для «будущих» (57%) и «прошлых» (55%). Именно такой результат и следовало ожидать на основе имеющихся теоретических представлений об эволюционной «гонке вооружений». Попробуем разобраться, почему.
Ранее было установлено, что устойчивость дафний к тем или иным штаммам бактерии Pasteuria ramosa предопределена генетически. У дафний имеется некий не очень большой набор генов, определяющих устойчивость к паразиту, причем каждый ген существует в виде нескольких вариантов (аллелей). Точно число этих генов и аллелей неизвестно, однако установлено, что в зависимости от конкретной комбинации аллелей рачок оказывается более или менее устойчивым к данному штамму бактерии. Популяция бактерий, со своей стороны, тоже генетически разнородна, причем разные линии (штаммы) паразита характеризуются разной инфекционностью по отношению к разным генетическим вариантам хозяина.
Возьмем простейший случай: допустим, что существует всего два генетических варианта дафний (А и Б) и всего две разновидности бактерий (А1 и Б1). Предположим, что бактерия А1 эффективно заражает дафний А, но не может заразить дафний Б, а бактерии Б1, напротив, успешно расправляются только с дафниями Б.
Допустим, исходно в пруду преобладали дафнии А. Но вот в пруд попадают бактерии-паразиты А1 и Б1. Первый из этих штаммов начинает быстро размножаться, поскольку для него имеется много «пищи» — дафний А. Складывается популяция бактерий с преобладанием варианта А1. Именно такая популяция бактерий наиболее опасна для тех дафний, которые сейчас живут в пруду, поскольку среди них преобладает вариант А, уязвимый для доминирующей разновидности паразита.
Известно, что бактерии Pasteuria снижают плодовитость зараженных дафний. Таким образом, под действием бактериальной инфекции в популяции дафний начинается естественный отбор: дафнии Б будут оставлять больше потомства, чем дафнии А. В результате через несколько поколений форма Б становится преобладающей среди дафний. Тем самым популяция дафний, по сути дела, приспособилась к инфекции. Для большинства дафний теперь не опасен наиболее массовый штамм бактерии (А1).
Но радоваться дафниям недолго. Произошедшие изменения в структуре популяции хозяев немедленно меняют вектор отбора, действующий на бактерий: теперь преимущество получают бактерии Б1, а доля бактерий А1 начинает снижаться. У бактерий смена поколений происходит гораздо быстрее, чем у дафний, и поэтому за очень небольшой отрезок времени популяция бактерий приобретает структуру, наиболее опасную для нынешней популяции дафний: теперь среди бактерий преобладает форма Б1, и дафниям приходится снова «приспосабливаться», то есть возвращаться в исходное состояние (когда преобладала форма А).
Таким образом, в системе будут происходить периодические колебания частот аллелей — бесконечное микроэволюционное состязание, напоминающее не столько гонку вооружений, сколько бег на месте. Несмотря на постоянно идущие быстрые изменения частот аллелей, суммарный итог этой борьбы скорее всего сведется к тому, что в каждый момент времени степень зараженности дафний и средняя инфекционность бактерий будут оставаться примерно одинаковыми, так что при поверхностном наблюдении может показаться, что система пребывает в состоянии полного покоя.
Следует помнить, что мы рассмотрели предельно упрощенный случай. В действительности генетических вариантов и у дафний, и у бактерий существует намного больше; могут появляться и новые варианты в результате мутаций, а у дафний существует еще и такая полезная вещь, как половое размножение, позволяющее резко увеличить генетическое разнообразие потомства. Дафнии, как известно, чередуют половое размножение с бесполым (партеногенетическим). В периоды партеногенетического размножения каждая самка производит потомство, генетически идентичное ей самой. В такие периоды паразитам намного легче «угнаться» за эволюционными изменениями хозяев. Ведь во время партеногенетического размножения эти изменения сводятся всего лишь к колебаниям частот уже имеющихся генетических комбинаций и редким мутациям. Переход к половому размножению означает внезапную перетасовку генетического материала хозяев. При этом возникает множество новых комбинаций аллелей, к которым паразиты еще не имели возможности приспособиться. Кстати, некоторые ученые предполагают, что само появление полового размножения на заре эволюции эукариот могло быть простимулировано именно такими «эволюционными гонками» в системах «паразит—хозяин».
Однако методика, использованная авторами обсуждаемой статьи, не позволяет проследить эволюционное соревнование во всех деталях, поскольку каждый исследованный слой осадка содержит «обобщенную» информацию за период длительностью в 2–4 года. За это время дафнии успевают несколько раз перейти от бесполого размножения к половому и обратно. По той же причине не удалось зафиксировать и фазовый сдвиг, который непременно должен присутствовать в циклической эволюционной динамике паразитов и хозяев. Бактерии, конечно, приспосабливаются к изменению частот аллелей у дафний очень быстро, но всё же не мгновенно. Поэтому, если бы была возможность проследить процесс поколение за поколением, то скорее всего можно было бы увидеть, что настройка параметров инфекционности у бактерий чуть-чуть отстает от настройки параметров защиты у дафний.
Иными словами, самыми опасными для дафний по идее должны являться не современные им популяции паразитов, а те, которые возникнут в недалеком будущем. Но, поскольку в данном исследовании фактически усреднялись данные за 10–20 поколений дафний, этот сдвиг зарегистрировать не удалось, и получилось, что самыми инфекционными являются «современные» популяции паразитов.
Возможность эволюционных «гонок на месте» впервые предположил в 1970-е годы американский палеонтолог Ли Ван Вален (Leigh M. Van Valen). Он назвал свою модель «гипотезой Черной Королевы» (The Red Queen Hypothesis). Имеется в виду персонаж из «Алисы в Зазеркалье» — Черная Королева, которая бежала со всех ног только для того, чтобы остаться на месте. (Для знатоков английского нужно пояснить, что в сказке Льюиса Кэрролла речь идет о шахматных фигурах, которые у нас — белые и черные, а у них — белые и красные, почему и приходится «Red Queen» переводить как «Черная Королева».)
Ван Вален хотел объяснить, почему вероятность вымирания видов не снижается с возрастом. Казалось бы, в результате естественного отбора «приспособленность» вида должна всё время повышаться, и поэтому старые виды должны быть более «приспособленными», чем молодые. Но это не подтверждается палеонтологическими данными. Ван Вален предположил, что почти все силы у эволюционирующего вида расходуются на то, чтобы угнаться за постоянно меняющейся внешней средой (в том числе за эволюцией других видов, с которыми данный вид взаимодействует). Вид всё время приспосабливается к среде, но из-за переменчивости условий его итоговая приспособленность в лучшем случае остается постоянной — вот и получается «бег на месте».
Гипотеза Ван Валена приобрела широкую популярность среди биологов самых разных специальностей, однако прямых экспериментальных подтверждений у нее пока немного. Поэтому данное исследование вносит весомый вклад в теоретическую биологию.
")
")