Сеть трофических (пищевых) взаимосвязей в природных экосистемах устроена асимметрично. Энергия в них передается от нижних уровней к верхним по разным каналам (один — более быстрый, другой — более медленный), которые объединяются хищниками только на самых верхних уровнях.
В середине ХХ века экологи, казалось, были полностью уверены в том, что чем сложнее устроена экосистема, чем больше в ней видов, тем более она стабильна. Такую точку зрения отстаивал в частности выдающийся британский эколог Чарлз Элтон (Charles Sutherland Elton) в своей книге «Экология нашествий животных и растений» (The Ecology of Invasions by Animals and Plants, английское издание 1958 года, русское 1960 года, последнее английское переиздание 2000 года). Однако в 1970-е годы Роберт Мэй (Lord (Robert) May of Oxford), уроженец Австралии, по образованию инженер, ставший физиком-теоретиком, а впоследствии экологом, показал на математических моделях, что в системе, которая состоит из множества элементов, связанных между собой отношениями типа «хищник—жертва», увеличение числа элементов само по себе не приводит к возрастанию стабильности. Скорее, наоборот: более сложные системы оказываются нестабильными. Однако, поскольку реальные природные экосистемы порой бывают весьма сложными и при этом устойчивыми, очевидно, что всё дело не просто в числе взаимодействующих элементов, а в том, как они соединены между собой, в особенностях трофической (пищевой) структуры системы.
|

Общим принципам организации трофических сетей посвящена большая статья, появившаяся в одном из последних номеров журнала Nature. Ее авторы — Найл Руни (Neil Rooney) из Отдела интегративной биологии Университета в Гуэлфе, Онтарио, Канада и его коллеги из того же университета и из Университета Северного Колорадо (США) — утверждают, что основной принцип устройства природных экосистем — это асимметрия. Проявляется она в том, что вещество (а с ним и энергия) в большинстве экосистем передаются от нижнего трофического уровня к верхнему по крайней мере двумя потоками («каналами»), различающимися по своей скорости и мощности. Слияние этих двух каналов происходит только на самых верхних уровнях, где располагаются так называемые «верховные хищники» (top predators).
В качестве примера можно привести систему трофических связей в озере. В ее основании фитопланктон (создающий органическое вещество, которое потом может быть использовано зоопланктоном) и детрит — взвешенное неживое органическое вещество, которое может быть использовано бактериями, в свою очередь потребляемыми некоторыми животными. Потребители фитопланктона и бактерий могут быть разными, но в составе рациона хищных рыб, находящихся на самом верхнем трофическом уровне, есть виды, представляющие как один, так и другой канал (см. также Прожить в озере, не питаясь едой с суши, нельзя). При этом надо иметь в виду, что хотя на каждом уровне организмы тратят немало энергии для поддержания своего существования и верховным хищникам остается ее совсем немного, роль последних в регуляции (сдерживании роста численности) своих жертв может быть определяющей.
Проанализировав подробные опубликованные данные по трофическим сетям четырех наземных и четырех водных экосистем, авторы в каждой из систем выделили по два «канала» передачи энергии и обнаружили, что по мере возрастания трофического статуса (положения в трофической цепи) того или иного вида (или группы видов) увеличивается их зависимость не от одного, а от двух каналов. Высота уровня может быть дробной величиной, поскольку она вычисляется по процентному соотношению энергии, получаемой данными организмами с разных уровней. Например, добычей крупных хищных рыб могут быть как более мелкие хищные рыбы, так и те рыбы, которые питаются планктоном, то есть находятся в трофической цепи на одну ступеньку ниже.
|
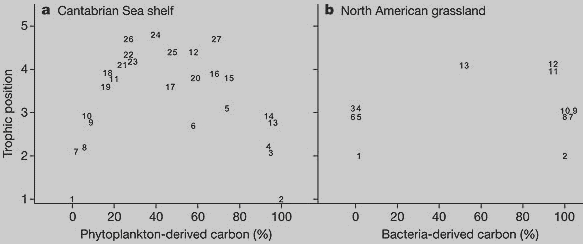
Выяснилось также, что «каналы», начинающиеся, к примеру, в почвенной экосистеме от грибов или от бактерий, а в водных экосистемах — от планктонных или бентосных (бентос — совокупность организмов, на дне и в грунте морских и материковых водоемов) организмов, различаются по скорости передачи энергии. Бактериальный и планктонный каналы быстрее, чем грибной и бентосный: они характеризуются преобладанием организмов с более высоким отношением продукции к биомассе.
Изучение поведения математических моделей таких систем показало, что в тех случаях, когда потоки по двум каналам уравниваются, система становится гораздо менее стабильной. Дело в том, что в этом случае изменения в обоих каналах происходят синхронно, а важна именно асинхронность и возможность компенсации одного за счет другого. Резкое снижение стабильности происходит и в том случае, когда поток идет только по одному каналу. Важное значение в стабилизации принадлежит «верховным хищникам», зависящим от обоих каналов и способных эффективно сдерживать рост тех жертв, которые начинают преобладать.
Отсюда следуют и важные практические выводы. Для сохранения своей устойчивости экосистемы должны быть гетерогенными, допускающими возможность разных путей («каналов») передачи вещества и энергии от основания трофической пирамиды до ее вершины. Кроме того, в них должны быть «верховные хищники», способные переключаться с одного канала на другой.
")
")