Одна из самых сложных задач в нейробиологии — установление связи между поведением и нейронной активностью. Понятно, что то или иное движение и комплекс движений будут сопровождаться специфической нейронной деятельностью в мозге, и такая активность при одном движения будет отличаться от активности при другом движении. Обычно, чтобы сопоставить нейронную работу с поведением, просто берут некий поведенческий акт и начинают изучать сопутствующую ему активность в мозге. Но это требует огромных трудозатрат, не говоря уже о том, что разные типы поведения могут комбинироваться друг с другом, переходить друг в друга и т. д.
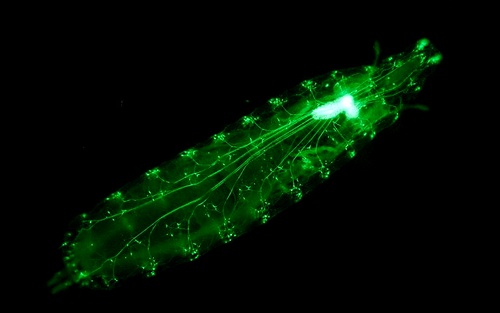
Совсем иной подход к этой задаче предлагают исследователи из Университета Джонса Хопкинса и Университета Дьюка (оба — США). Для начала Джошуа Вогельштейн (Joshua Vogelstein) и его коллеги получили генетически модифицированных личинок дрозофил, у которых в нейронах можно было включать синтез фоточувствительного белка: после того как этот белок появлялся, нейроны можно было активировать светом. Сейчас такие оптогенетические методы используются весьма широко, и как раз в нейробиологических исследованиях. Однако в этой работе была одна особенность: исследователи промодифицировали свыше тысячи нейронных цепей, для этого им потребовалось цветы купить.
Если простимулировать нейрон, это вызовет реакцию: организм совершит некое движение, куда-нибудь поползёт и т. д. Именно этим авторы работы потом и занялись: они стимулировали светом модифицированные нейроны у личинок мух и с помощью специального оборудования, фиксировавшего любое движение насекомых, наблюдали за их поведением. В итоге на руках оказался огромный массив данных — записи движений 37 780 личинок.
Разбираться со всем этим пришлось математическим статистикам. Вместе с биологами они сумели распределить элементарные движения личинок по нескольким типам поведения. Таких типов, которые включали в себя последовательности движений, оказалось 29, причём надо понимать, что выглядело всё это как ветвящееся дерево, ветки которого — разные типы поведения — выходили одна из другой, так как некоторые движения были общими для разных типов.
Среди поведенческих схем были довольно очевидные и легко отличимые друг от друга, которые можно было обозначить условно как «возвращение» или «безоглядное бегство». Однако наряду с такими типами обнаружились и другие модели поведения, которые до сих пор для мушиных личинок (а это всё-таки известный модельный объект в биологии) никто не описывал. Наконец, последовательности двигательных реакций были соотнесены с группами нейронов. Именно то, что авторы работали с группами клеток, позволяет говорить о надёжности результатов: группа нейронов с куда большей вероятностью специфично относится к определённому типу поведения, нежели пара–тройка клеток.
Конечно, насчёт поведения, на которое раньше не обращали внимания, можно сказать, что тут сработал фактор исследователя, провоцирующего организм на несвойственные ему действия. Однако целью работы было не столько описание естественного поведения личинок дрозофил, сколько создание метода, алгоритма, который позволял бы наносить разные варианты поведения на карту нейронных сетей. То есть теперь, видя, как личинка дрозофилы изгибается и ползёт к еде, можно с уверенностью сказать, какие клеточные кластеры у неё в мозге в этот момент работают, как именно они работают, как они сменяют друг друга.
Большой плюс метода, конечно, в том, что, несмотря на устрашающий объём данных, он позволяет получить намного больше информации за куда меньшее время, чем если бы мы разглядывали поведенческие акты, что называется, «поштучно». И, возможно, в скором времени такие подробные нейронные расшифровки появятся и для других животных, вплоть до шимпанзе.

")