
Самым "убедительным" примером промежуточного звена между двумя большими группами животных общепризнана "праптица" – археоптерикс.
Археоптерикс. С 1861 года было найдено в Золнхофенских плиточных известняках наряду с одним отдельным пером пять более или менее хорошо сохранившихся окаменевших экземпляров этих птиц, к которым добавилась еще и недавняя шестая находка. На них явно просматриваются как признаки пресмыкающихся, так и признаки птиц Типичными признаками рептилий считаются заполненная зубами челюсть, отсутствие киля на грудине, строение когтей и позвонков, свободные ребра без крюкообразных отростков и удлиненный хвостовой отдел позвоночного столба. Им противопоставлены типичные признаки птиц, такие как оперение, скелет рук, таза и ног.
Из-за их чрезвычайной значимости ископаемые археоптериксы подверглись тщательному и всестороннему изучению. Полученные при этом результаты привели к достаточно разным, отчасти даже противоречащим интерпретациям. Так, например, до настоящего времени дискутируется вопрос, следует ли считать археоптерикса прямым предком птиц (к чему до недавнего времени склонялось большинство исследователей) или формой, которая вела в эволюционный тупик. Характерным признаком птиц являются перья. Окаменелость, которая интерпретируется как предшественник птиц, должна была, вероятно, отличаться наличием "праперьев", т.е. образований, которые послужили бы перемычкой в процессе превращения, развития перьев из чешуи рептилий. Но как раз этого с археоптериксами и не происходит. Напротив, перья "праптиц" до микроскопических деталей повторяют строение перьев современных птиц.
К тому же имеются ископаемые находки фрагментов перьев из Бернштейна, низов нижнемелового периода (т. е. слоев, которые откладывались непосредственно над верхнеюрскими слоями). Консервация в Бернштейне позволяет производить микроскопические исследования, используя всю разрешающую способность светового микроскопа. При этом проявляется полное соответствие тонкой структуры перьев ископаемого экземпляра и перьев современных птиц. Так как подобные ископаемые перья были найдены и в Австралии и в Ливии, кроме того, отпечатки перьев встречаются в сланцах нижнемелового периода в Сибири и Монголии, а отпечатки птичьих лап – в нижнемеловом периоде в Канаде,’ то это должно было бы означать, что уже тогда птицы, несмотря на приписываемую им плохую способность летать, были распространены по всему миру.

В Монголии палеонтологи обнаружили, кроме упомянутых отпечатков перьев (в тех же самых слоях), останки птицы с четко обозначенным грудинным килем. Этот образованный на грудине костяной гребень служит местом фиксации мускулатуры, обеспечивающей полет, и рассматривается как типичный орган для "высокоразвитых" видов птиц, обладающих хорошо развитой способностью летать. Т. о., подобные виды птиц должны были появиться сразу же после археоптериксов. Все сказанное выше касается и находки фламингоподобных птиц из самых низов нижнемелового периода во Франции. Из отложений мелового периода всемирно известны также окаменевшие пингвины. Согласно эволюционной модели, эти данные свидетельствуют о том, что у некоторых видов птиц способность летать претерпела обратное развитие (атрофировалась) практически сразу после ее появления. (При этом остается открытым вопрос, как могло возникнуть необходимое для этого давление отбора взаимоисключающей направленности).
Упомянутые признаки пресмыкающихся у археоптериксов должны помочь найти прямых "предков первобытных птиц" в классе рептилий или, по крайней мере, помочь разыскать группы рептилий, из которых могли бы произойти эти "праптицы". При этом, однако, возникают большие проблемы, так как отдельные присущие археоптериксу признаки можно встретить у абсолютно разных групп рептилий, взаимоисключающих друг друга в качестве предшественников "праптицы": вид зубов
археоптерикса говорит, например, в пользу того, что его прямым предком является крокодил (крокодилообразные ящерицы). Археоптерикс обладает типичным признаком, присущим птицам, – вилочкой, которая образовало, из сросшихся костей ключицы (но многие попугаи и туканы имеют только ключицы, так неужели это может дать основание считать их более древними?). Так как множество амфибий имеют ключицы, то не следует ли провести прямую линию от земноводных к птицам? Однако эта линия должна миновать крокодилов, так как они не имеют ключицы. Кроме того, существуют значительные различия в строении тазобедренного пояса. Несмотря на то, что именно крокодилы имеют соответствующей формы зубы (хотя некоторые авторы оспаривают это), а также сходное строение слухового аппарата и челюстного сустава, различия в строении других органов перевешивают это сходство и практически исключают их из возможных претендентов на роль предшественников птиц. Едва ли кто-нибудь сегодня считает, что археоптерикс произошел от этого отряда рептилий.
Наличие зубов могло бы как раз служить доказательством его происхождения от рептилий, но скорее этот факт свидетельствует о том, что когда-то существовали птицы, которые имели зубы, но впоследствии вымерли (для сравнения вспомните о птицах верхнемелового периода, имевших зубы, которые, однако, по мнению ученых КУНА ШИНДЛЕРА и РИЕБЕРА (1984, с. 242), никак не могли иметь своим предком археоптерикса). У всех групп позвоночных животных встречаются виды, имеющие зубы и не имеющие их!
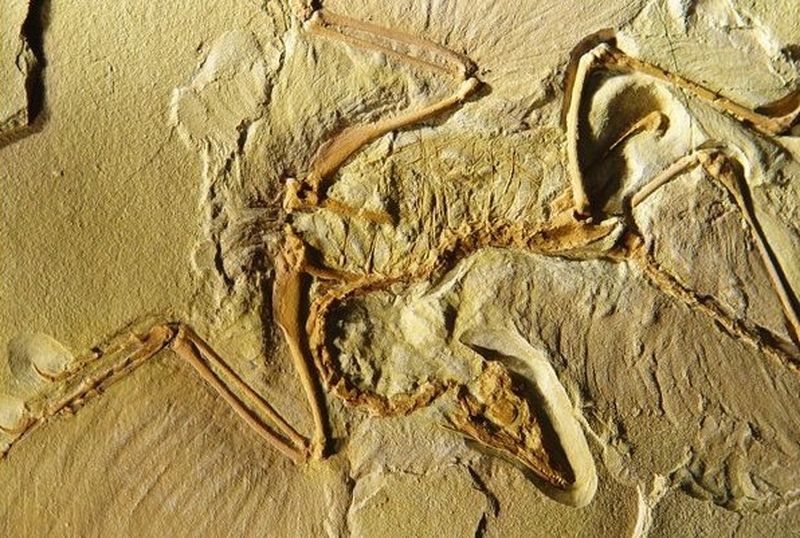
Археоптерикс обладает амфицельными позвонками (вогнутыми с обеих торцевых поверхностей). Однако подобный тип позвонков не встречается ни у современных, ни у ископаемых птиц. Он типичен для ихтиозавров, которые никогда и не рассматривались в качестве предшественников древних птиц. Тазобедренный пояс археоптерикса имеет четырехлучевое строение, сходное со строением орнитисхий (птицеящеров), одного из двух отрядов динозавров. Но и эта группа не имеет ключиц, что затрудняет возможность выведения происхождения птиц от этих рептилий. И хотя орнитисхий обладают сходным с птицами тазобедренным поясом, вряд ли от них можно вывести происхождение археоптериксов.
Когти на передних конечностях археоптерикса имеют большое сходство с подобными же рода Qmitholestes, который относится ко второму отряду динозавров – Saurischia. Но в качестве предшественников они тоже не рассматриваются, так как имеют трехлучевое строение таза. Во всяком случае, встречаются палеонтологи, которые реконструируют строение таза археоптерикса похожим на строение рода Соmpsognathus – маленьких динозавров, обнаруженных в Золнхофенских плиточных известняках. Т.о., вид Compsognathus является "современником" археоптерикса, а потому не может рассматриваться в качестве его "предшественника" (STEPHAN, 1987, с. 95). Подвижные когтевые фаланги можно встретить также и у современных птиц (например, гоацин). Как и у них, у археоптерикса были подвижными только когтевые фаланги, а не пальцы, так как на них и у "праптиц" размещались маховые перья. Что касается строения запястья, то, судя по нему, гоацин скорее похож на рептилий, чем на археоптериксов.
Вообще говоря, у археоптерикса, кажется, многие признаки, свойственные рептилиям, те же, что и у легких динозавров – тероподов, от которых он и происходит, несмотря на выше упомянутые возражения большинства (но не всех) современных палеонтологов. Комбинация множества признаков из различных групп пресмыкающихся в археоптериксе свидетельствует о том, что он является типичной мозаичной формой, и его нельзя больше рассматривать в качестве предка "современных" птиц. Мозаичные типы широко распространены в мире животных.
Обнаруженная недавно в Испании ископаемая птица нижнемелового периода проявляет себя как типичная мозаичная форма. Так называемая "птица из Лас-Хойас" демонстрирует, кроме хорошо сохранившихся перьев, комбинацию "примитивных" (древних) и "производных" (современных) признаков. К первым относится форма тазобедренного пояса, крестцовый позвоночник и скелет ноги, к последним – форма коракоида, и сросшийся хвостовой отдел позвоночника. В то время как формирование подвздошной кости (отдела таза, см. рис. 8.68) у "птицы из Лас-Хойас", согласно эволюционной модели, должно интерпретироваться как "более примитивное", форма коракоида соответствует ему же у современных Carinatae (птицы, имеющие килевидную, грудину). "Птица из Лас-Хойас" не может классифицироваться как недостающее звено между археоптериксом и современными птицами, но ее существование подтверждает лишь частое появление мозаичных форм.
Типичные признаки, присущие птицам, можно найти и у рептилий. Так, например, крокодилы обладают полной сердечной перегородкой и частично мускулистым желудком. Высиживающий потомство из яиц питон вырабатывает собственное тепло; и хотя он является холоднокровным", питон сопревает свою кладку тем, что обнимает ее своим телом. Плащеносная ящерица, обитающая в Австралии, время от времени ходит вертикально, опираясь на задние конечности.
Особенное строение скелета "древней птицы" не может здесь обсуждаться в полном объеме, приведем лишь еще несколько доказательств. Свободные ребра без крючкообразных отростков можно встретить и в современном мире птиц, даже у паламодей (Anhimidae). Следующим довольно часто упоминаемым "примитивным признаком" археоптерикса является отсутствие киля на грудине. Но он отсутствует почти полностью и у новозеландских совиных попугаев, а у большинства буревестников киль развит очень мало. В пользу подобного образа жизни мог бы говорить вытянутый костистый хвостовой отдел позвоночного столба, пожалуй, самый необычный признак археоптерикса. Возможно, именно длинный и широкий хвост делал его таким хорошим планеристом. (Так, например, коршуны, как хорошие планеристы, имеют вместо 12 рулевых перьев – 14, как у всех хищных птиц). Зачатки хвостового отдела в позвоночном столбе в период эмбрионального развития современных птиц встречается только у Carinatae ("современных птиц"), а не у предположительно древних по происхождению Ratitae, так что этот аргумент не может употребляться в рамках "биогенетического основного правила".
О летательных способностях археоптерикса можно было бы с уверенностью сказать, только если иметь четкое представление о состоянии атмосферы того времени. Возможно, археоптерикс был "только" планеристом, и это было его экологической нишей, которая ныне более не существует, так как изменилась плотность атмосферы. Если плотность атмосферы раньше была выше, то вполне вероятно, что "праптицы" могли быть не только хорошими планеристами, но и активно летали (несмотря на отсутствие киля на грудине). Доказательством того, что в древности существовали другие атмосферные условия, являются гигантские формы летающих ящеров (размах крыльев около 15 м!), птиц (хищные птицы Argentarius из североамериканского миоцена, имевшие размах крыльев около 7,5м) и насекомых (например, стрекозы). Есть все основания предполагать, что при сегодняшних атмосферных условиях летающие ящеры с размахом крыльев более 8 метров вряд ли были бы способны активно летать.
Для более точной классификации археоптерикса в постулированном родословном дереве птиц обратимся к специалистам-биологам. Так, например, Д. С. ПЕТЕРС и В. Ф. ГУГМАНН пишут: "Если рассматривать археоптерикса вне теоретического контекста (эволюционной теории, прим. авторов), то он не что иное, как необычная окаменелость" (1976, с. 265). Е. БЕЦЦЕЛЬ пишет в своей книге "Орнитология" (1977, с. 230): "Археоптерикс в классификационной системе стоит относительно изолированно, так как нам до сих пор неизвестны ископаемые, которые, с одной стороны, указывали бы на его связь с предками-рептилиями, а с другой стороны, устанавливали бы его связь с современными птицами". И еще: "До настоящего времени не выяснено, был ли, вообще, археоптерикс в момент начала развития других птиц" (с. 231). Если судить по наличию перьев, то археоптерикса можно с уверенностью отнести к птицам. Как мозаичный тип, он стоит абсолютно изолированно, так как до сих пор не найдено подобных видов. В качестве переходной формы между пресмыкающимися и птицами его, однако, рассматривать нельзя. Его нельзя считать таковым уже хотя бы потому, что в породах позднейшего нижнемелового периода, без сомнений, встречаются окаменевшие останки "современных" птиц.
Недавно (1986) палеонтолог С. ЧАТТЕРЖИ (Технический университет, Техас) дал следующее разъяснение ископаемым костям, которые были открыты в • Техасе в породах, которые были датированы возрастом в 225 миллионов лет: это останки пернатых животных, способных летать. Названный им Protoavis ("первоптица") вид животных должен был обладать способностью перелетать с дерева на дерево. Среди находок встречаются полые кости, малый вес которых позволяет птицам летать, коническая грудная кость, такая же, как у современных птиц, и кость черепа, имеющая форму, подобную птичьей. Если считать толкование этой ископаемой находки, данной С. ЧАТТЕРЖИ, удовлетворительным (хотя эти выводы оспариваются другими палеонтологами (см, ОСТРОМ, 1987), то археоптерикс должен был бы занять свое место в тупиковой побочной ветви эволюционного развития, так как, согласно традиционной датировке, археоптерикс, по крайней мере, на 75 миллионов лет должен быть моложе, чем Protoavis.
В результате подробного изучения специалистами археоптерикс определяется скорее как вымершая "побочная ветвь" в ходе эволюции птиц, а в эволюционно-теоретических рамках – в лучшем случае, как "модель" промежуточной ступени. Кроме всего прочего, ископаемые археоптериксы имеют огромное значение, так как они дают нам представление о своеобразном вымершем типе птиц.
С 1984 года в международной прессе появляются все новые публикации о том, что археоптериксы – это подтасовка ископаемых данных. Приводятся аргументы в пользу этой позиции; по мнению авторов, наиболее убедительно это доказывают ученые РИЧЕЛ (1985, с. 371) и ЧАРИЖ с группой ученых (1986).
")
")