Компьютерное моделирование показало, что вероятность симпатрического видообразования (разделения исходного вида на два в отсутствие географических барьеров) резко повышается, если у самок есть возможность выбирать партнеров на основе признаков, свидетельствующих о хорошем здоровье самца. Как ни странно, половой отбор резко ускоряет видообразование даже в том случае, если самки двух зарождающихся видов используют при выборе самцов одни и те же критерии.
Всего пару десятилетий назад симпатрическое видообразование считалось крайне маловероятным или вообще невозможным. Однако целый ряд новых блестящих открытий показал, что это не только вполне реальное, но и весьма распространенное в природе явление (список хорошо изученных случаев симпатрического видообразования и подборка ссылок по теме приведены в заметке Зафиксирован начальный этап видообразования у тропических бабочек, «Элементы», 09.11.2009).
В изучении симпатрического видообразования теория несколько отстала от практики: конкретные случаи удалось зарегистрировать и изучить, но с механизмами, заставляющими единую популяцию разделиться на две репродуктивно изолированные части, пока еще нет полной ясности. Чтобы симпатрическое видообразование окончательно утвердилось в науке в качестве «полноправного» эволюционного механизма, необходимы хорошие математические модели, показывающие, как «устроен» этот процесс и от каких факторов он зависит.
В истории эволюционной биологии не раз бывало так, что хорошая идея, давно уже высказанная каким-нибудь гениальным теоретиком и даже подкрепленная фактами и наблюдениями, долго оставалась «в тени» и не находила всеобщего признания до тех пор, пока кто-нибудь не разрабатывал математическую модель, которая объяснила бы всем, как и почему эта идея работает. Больше всех не повезло теории полового отбора. Эта абсолютно гениальная (как мы теперь понимаем) идея была подробно разработана Дарвином (см.: Ч. Дарвин. Происхождение человека и половой отбор; Djvu, 20 Мб), но современники ее не поняли и не приняли. Ученые просто не могли поверить, что самки могут активно выбирать самцов. Самки, по мнению мыслителей Викторианской эпохи, могут лишь скромно сидеть в уголочке в нарядном платье и ждать, пока к ним кто-то посватается.
Идея полового отбора успела стать «полузабытым научным курьезом», когда в 1930 году Рональд Фишер эксгумировал ее, развил и дополнил важными деталями, до которых Дарвин не додумался (см.: R. Fisher. The Genetical Theory of Natural Selection). В частности, Фишер понял, что не только брачные украшения самцов, но и вкусы и избирательность самок тоже наследуются и эволюционируют по тем же правилам, что и остальные признаки. Эта простая мысль заткнула самые главные прорехи в теории Дарвина и сделала ее полностью работоспособной. Сегодня это кажется невероятным, но идеи Фишера о половом отборе тоже не были восприняты научным сообществом. Они почти забылись к тому времени, когда их снова выкопали из забвения представители следующего поколения биологов-теоретиков, сумевшие, наконец, разработать убедительные математические модели. Лишь с третьей попытки теория полового отбора наконец получила заслуженное признание. Эта драматическая история красочно изложена в книге Geoffrey Miller. The Mating Mind: How Sexual Choice Shaped the Evolution of Human Nature (PDF, 2,1 Мб).
Сегодня уже никто не сомневается, что половой отбор — мощнейшая эволюционная сила, способная многократно усиливать эффективность «обычного» естественного отбора и ускорять адаптивную эволюцию (а не только создавать «бессмысленные», ненужные для выживания украшения и причудливое брачное поведение). Если самки выбирают самцов не по произвольным бессмысленным критериям, а по признакам, отражающим приспособленность или «качество генов» самца, — например, по признакам, свидетельствующим о здоровье и хорошей физической форме, — то тем самым самки резко ускоряют эволюцию.
Легко понять, почему половой отбор повышает эффективность естественного отбора: самец с пониженной приспособленностью не только имеет меньше шансов выжить, но и становится менее привлекательным для самок. Даже очень небольшие различия в приспособленности, едва заметные для «обычного» отбора, могут стать решающими, когда дело доходит до конкуренции между самцами в попытках очаровать привередливую самку. Кроме того, отбор перестает быть «слепым». Теперь его направляют существа, у которых всё же есть кое-какие мозги. Эти существа способны к целенаправленным действиям и отчасти осмысленным решениям. Самки становятся селекционерами. Привередливые павлинихи создали роскошные узоры на хвостах самцов-павлинов точно так же, как голубеводы создали причудливые украшения у декоративных пород голубей.
Прочно закрепившись в науке, теория полового отбора сегодня помогает вытащить из тени другую важную и незаслуженно забытую идею — симпатрическое видообразование. Многим теоретикам давно было ясно, что без полового отбора и избирательного скрещивания симпатрическое видообразование едва ли возможно. Проблема была в том, что всем казалось, будто для симпатрического видообразования самки двух зарождающихся видов должны иметь разные половые предпочтения. И было не очень понятно, с чего бы вдруг у самок в одной популяции возникли разные вкусы (см.: Как отличить своих от чужих? Неканонические механизмы репродуктивной изоляции).
Сандер ван Дорн из Бернского университета (Швейцария) и его коллеги из Гронингенского университета (Нидерланды) опубликовали в последнем номере журнала Science статью, которая может окончательно превратить симпатрическое видообразование из маргинальной идеи в общепринятую теорию. Размах чувствуется уже в названии статьи. Без ложной скромности авторы озаглавили ее «О происхождении видов путем естественного и полового отбора», что является точным повторением заглавия перевернувшей мир книги Чарльза Дарвина, за исключением предпоследнего слова.
Модель ван Дорна и его коллег показывает, что даже если все самки в популяции имеют одинаковые половые предпочтения, их привередливость может радикально повысить вероятность симпатрического видообразования. Разумеется, для этого должен выполняться некий минимум условий.
Всеми признано, что половой отбор по признакам, свидетельствующим о хорошем здоровье или хороших генах (такие признаки называют «индикаторами приспособленности») повышает эффективность, во-первых, негативного (очищающего, стабилизирующего) отбора, то есть отбраковки вредных мутаций, во-вторых — позитивного (движущего) отбора, то есть фиксации полезных мутаций. Основная идея ван Дорна и его коллег, воплощенная в модели, состоит в том, что отбор по «индикаторам приспособленности» повышает также и эффективность дизруптивного (разрывающего, разводящего) отбора. Дизруптивный отбор действует в том случае, если крайние варианты какого-то признака более адаптивны (выгодны), чем промежуточные.
|
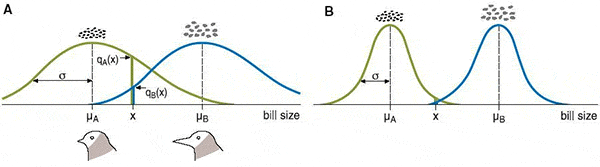
Допустим, существует популяция птиц с клювами средней длины. Местность, где они живут, состоит из участков двух типов. На одних участках преобладают растения с крупными семенами, на других — с мелкими. Для разгрызания крупных семян удобнее иметь длинный клюв, для мелких — короткий. В такой ситуации может начаться дизруптивный отбор. Это значит, что преимущество получат птицы с более длинными или более короткими клювами, но не с промежуточными. Известно, что у реальных птиц, попавших в подобные условия, может произойти симпатрическое видообразование (см.: Видообразование на разных островах идет параллельными путями, «Элементы», 15.03.2007). Остается только понять, каким образом видам удается разделиться, если их представители, перелетая с участка на участок, то и дело встречаются друг с другом и никто не мешает им образовывать смешанные пары и производить гибридное потомство с промежуточными по длине клювами.
|
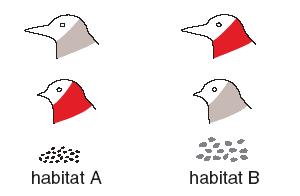
В рассматриваемой модели все птицы изначально имеют одинаковые клювы средней длины, у самцов нет «индикаторов приспособленности», а у самок — избирательности при выборе партнера, поэтому скрещивания происходят случайным образом. Все признаки могут меняться в результате мутаций. Для большего правдоподобия длина клюва зависит не от одного, а от десяти генов. Взрослые птицы живут на своих участках, молодые могут мигрировать на другие участки. Специальная переменная задает интенсивность миграций. На участках с крупными семенами лучше себя чувствуют особи с большими клювами, и наоборот. Птица, которая лучше питается, имеет больше шансов дожить до зрелости. Кроме того, ее организм располагает большим количеством ресурсов. Самцы могут направить часть этих ресурсов на формирование красного оперения, которое служит «индикатором приспособленности». Самки могут приобрести способность выбирать самцов по этому признаку.
И красное оперение, и избирательность самок — «дорогостоящие» признаки. Это значит, что на их развитие тратится часть ресурсов организма, которые могли быть использованы на повышение выживаемости. Самцы платят за свою красоту снижением шансов на выживание. Именно поэтому данный признак может служить индикатором приспособленности. Если бы красное оперение было «бесплатным», все самцы быстро стали бы профессиональными обманщиками, а избирательность самок потеряла смысл. То, что избирательность самок тоже не является «бесплатным» признаком, менее принципиально, зато реалистично. Эволюция, как правило, пытается сэкономить на мозгах (см.: Обнаружить птенца-подкидыша — слишком сложная задача для птичьих мозгов?, «Элементы», 21.12.2009). Таким образом, в этой модели самцы не могут выработать других украшений, кроме красных перьев на шее. Самки, со своей стороны, не могут выработать других предпочтений, кроме любви к красношеим самцам. В частности, самки не могут научиться выбирать самцов по длине их клюва.
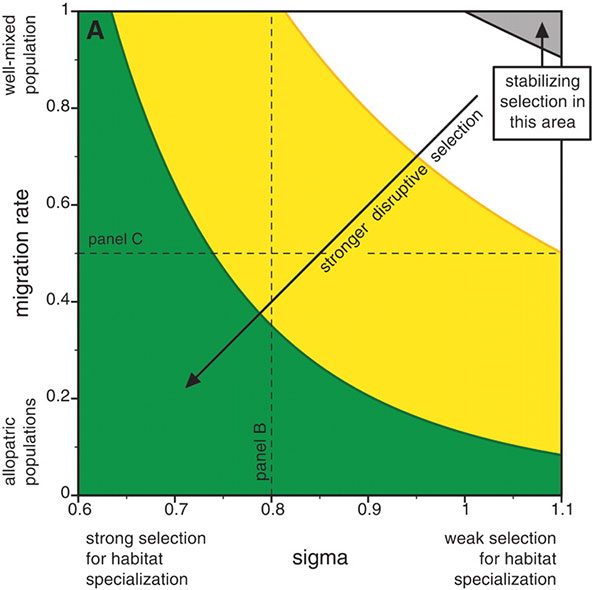
Главными входными параметрами модели являются интенсивность миграций молодых птиц и ширина кривой приспособленности (величина ? на рисунке). Чем больше сигма, тем легче разгрызать семена неподходящим по размеру клювом. Вместе эти две величины задают интенсивность дизруптивного отбора.
В этой модели у птиц нет возможности выработать репродуктивную изоляцию, то есть перестать скрещиваться с «чужаками» и производить гибридное потомство. Парадоксальным образом симпатрическое видообразование происходит и так. Под успешным видообразованием в данном случае понимается разделение популяции на две четко различающиеся по длине клюва субпопуляции с нулевой или крайне низкой численностью «промежуточных» особей.
|
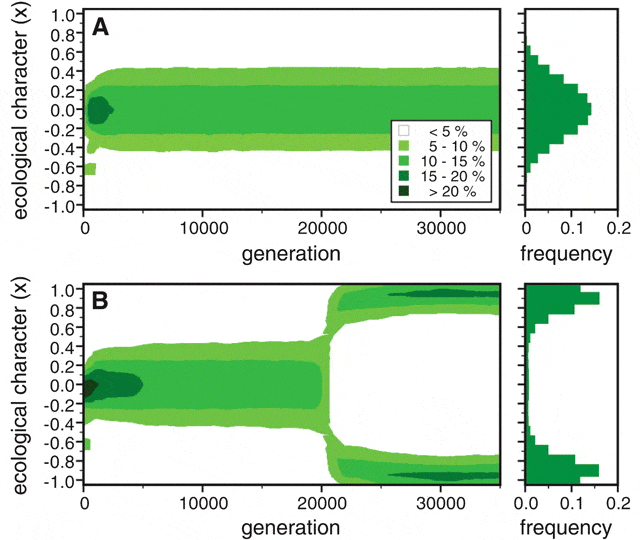
Сначала авторы исследовали поведение модели без «индикатора приспособленности» и половой избирательности. Как и следовало ожидать, в такой ситуации симпатрическое видообразование происходит только при экстремальных значениях параметров: либо при почти полном отсутствии миграций, либо при очень маленькой сигме. В остальных случаях силы дизруптивного отбора хватает лишь на то, чтобы поддерживать в популяции высокий уровень генетического полиморфизма и изменчивости по длине клюва, но «разорвать» популяцию надвое дизруптивному отбору не удается.
Затем модельным птицам «разрешили» приобретать в результате мутаций половую избирательность и способность расходовать часть ресурсов на красные перышки. Эффект оказался неожиданно сильным: теперь исходный вид стал распадаться надвое даже при активной миграции и большой сигме (рис. 4). Таким образом, модель показала, что половой отбор по индикатору приспособленности способен радикально повысить вероятность симпатрического видообразования.
|
На первый взгляд это кажется очень странным. Допустим, два вида уже разделились. Но при высоком уровне миграции многие короткоклювые самки будут селиться на участках с большими семенами. Если они выживут, то самцов будут выбирать по красному оперению. На участке с большими семенами красное оперение будет у самцов с большими клювами. Таким образом, короткоклювые самки на «неподходящем» для них участке систематически будут делать «неправильный» выбор и производить потомство с промежуточным размером клюва. Однако на рис. 3B мы видим, что частота встречаемости промежуточных клювов после успешного симпатрического видообразования близка к нулю. Как это получается?
Разгадка состоит в том, что приспособленность в этой модели, как и в жизни, — понятие относительное. Выживет ли на данном участке птица с данным размером клюва — это зависит не только (и не столько) от того, насколько ее клюв соответствует семенам на участке. Судьба птицы определяется исходом ее конкуренции с себе подобными. Если семена большие, а клюв у птицы маленький или средний, это еще не смертный приговор. Если у всех птиц вокруг тоже маленькие или средние клювы, у данной птицы есть хороший шанс выжить. Но если все кругом имеют большие клювы, ее шансы ничтожны. Любая популяция производит больше потомков, чем может прокормиться на данной территории. Выживают не те, кто «вообще» хорошо приспособлен к данной среде, а те, кто приспособлен хорошо по сравнению с остальными.
Таким образом, в исходной ситуации, когда у всех птиц средние по размеру клювы, на любом из участков можно нормально жить, имея средний клюв. Но как только популяция начинает распадаться на две, для среднеклювых птиц наступают тяжелые времена. На обоих типах участков их безжалостно вытесняют специалисты — длинноклювые и корткоклювые конкуренты. Половой отбор действует просто-напросто как усилитель дизруптивного отбора. Но как только популяции начинают расходиться, дизруптивный отбор усиливается дополнительно еще и из-за самого факта расхождения. Чем дальше оно зашло, тем эффективнее отбраковка промежуточных особей. В итоге отбор начинает убирать среднеклювых птиц из популяции быстрее, чем они нарождаются.
Разумеется, рассмотренный в данной модели механизм — не единственный возможный «катализатор» симпатрического видообразования. Симпатрическое видообразование станет еще более вероятным, если «разрешить» птицам в ходе эволюции менять алгоритм миграций. Например, птицы могут выработать алгоритм «мигрируй, только если вокруг нет удобных для тебя семян» или «селись только там, где много удобных семян». Это приведет к резкому снижению частоты гибридизации. Еще одна возможность связана с развитием механизма выбора партнера «с оглядкой на себя» или с использованием импринтинга (запечатления). Например, самка может предпочитать партнеров с таким же клювом, как у нее самой или у ее отца. Подобные примеры известны, в том числе и среди птиц (см.: Пол птенцов зависит от мнения самки о своем муже, «Элементы», 23.03.2009).

")