|

К билатериям относятся все животные, кроме самых примитивных (таких как губки, трихоплакс и кишечнополостные), в том числе все черви, моллюски, членистоногие и хордовые. Специалисты по сравнительной анатомии и эмбриологии в течение прошлого века разработали несколько альтернативных теорий происхождения и ранней эволюции билатерий. Решить, какая из этих моделей ближе всего к реальности, помогли новые данные, в том числе молекулярно-генетические. (Об этом подробно рассказано в статье В. В. Малахова «Новый взгляд на происхождение билатерий»).
|
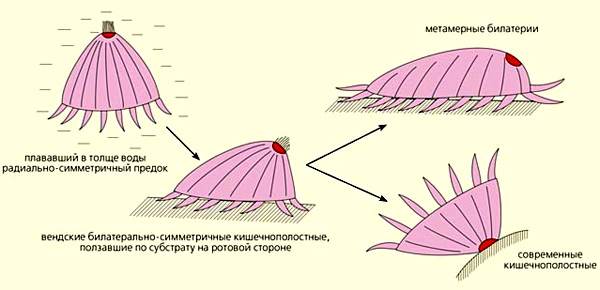
По современным представлениям, последний общий предок всех современных билатерий был сегментированным животным, имел вторичную полость тела (целом) и, возможно, парные конечности (параподии). Иными словами, из всех современных животных он был больше всего похож на кольчатого червя. Соответственно, кольчатые черви — наименее изменившиеся из всех его потомков, или, что то же самое, самые примитивные из современных билатерий. Это звучит непривычно для многих зоологов, которые привыкли считать самыми примитивными билатериями не кольчатых, а плоских червей. Однако филогенетические реконструкции, основанные на молекулярных данных, свидетельствуют о том, что плоские и круглые черви, устроенные проще кольчатых, произошли от более высокоорганизованных предков, то есть подверглись вторичному упрощению (подробнее см. в заметках: Конечности у членистоногих и кольчатых червей растут из общего корня, «Элементы», 20.11.2008; Новые данные позволили уточнить родословную животного царства, «Элементы», 10.04.2008).
Подобно тому как ланцетник может служить своего рода «живой упрощенной схемой» хордовых (см.: Геном ланцетника помог раскрыть секрет эволюционного успеха позвоночных, «Элементы», 23.06.2008), а изучение воротничковых жгутиконосцев помогает проникнуть в тайну происхождения первых многоклеточных животных (см.: Расшифрован геном хоанофлагеллят — ближайших одноклеточных родичей всех многоклеточных животных, «Элементы», 18.02.2008), кольчатые черви являются удобной моделью для изучения древнейших этапов эволюции билатерий.
Ранняя эволюция билатерий представляет особый интерес в связи с тем, что именно на этом этапе развития животного царства произошло радикальное усложнение и диверсификация планов строения и появилось много новых тканей и органов, в том числе центральная нервная система, разнообразные органы чувств, сквозной кишечник, специализированная мускулатура и т. д.
Чтобы понять, откуда взялись все эти новшества, необходимо прежде всего разобраться, как они возникают в ходе индивидуального развития (онтогенеза) современных организмов. Дело тут не только в том, что индивидуальное развитие иногда отчасти воспроизводит («рекапитулирует») эволюционную историю, но и в том, что эволюция строения организма — это прежде всего эволюция программы индивидуального развития. Ведь в геноме нет ничего похожего на «чертеж» взрослого животного. Там есть только «программа поведения», работающая на уровне отдельных клеток развивающегося эмбриона. Морфология взрослой особи является результатом процессов самоорганизации, основанных на согласованном поведении делящихся эмбриональных клеток. Иными словами, вопрос о том, как появились новые органы и ткани, сводится к вопросу об изменениях в программе онтогенеза, которые произошли у древних билатерий.
Важные шаги к пониманию этих изменений были сделаны в ходе изучения Hox-генов и других генов — регуляторов индивидуального развития животных (см.: Новое в науке о знаменитых Hox-генах, регуляторах развития, «Элементы», 10.10.2006). Стало ясно, что белки, кодируемые этими генами, играют важную роль в онтогенезе билатерий. Их наиболее очевидная функция состоит в том, что они «размечают» эмбрион, определяя, где будет голова, а где хвост, где спина, а где брюхо, где расти ногам, а где быть границам между сегментами. Манипулируя работой белков-регуляторов, можно превратить, например, третий сегмент груди дрозофилы в дубликат второго — и тогда у мухи вырастет лишняя пара крыльев, можно превратить антенны в ноги или добиться появления зачатков конечностей на брюшных сегментах, где им быть не положено.
Однако до понимания всех тонкостей программы онтогенеза билатерий нам по-прежнему до обидного далеко. В частности, выяснилось, что онтогенезом управляют не только белки, но и особые маленькие молекулы РНК — микроРНК (microRNA). Как и белки-регуляторы, микроРНК влияют на активность генов, причем под их контролем находятся также и гены, кодирующие те самые белки-регуляторы. Во многих случаях удалось установить, какие именно гены регулируются теми или иными микроРНК, но как всё это связано с конкретными планами строения билатерий и с их эволюцией, выяснить оказалось не так-то просто.
В новой статье германских ученых, опубликованной на сайте журнала Nature, предпринята попытка внести хоть какую-то ясность в вопрос о роли микроРНК в ранней эволюции билатерий. Для этого авторы изучили пространственное распределение разных микроРНК в органах и тканях развивающейся личинки кольчатого червя Platynereis, который по своему строению очень близок к общему предку билатерий, каким он видится сегодня биологам. Для сравнения использовался еще один кольчатый червь (Capitella) и морской еж Strongylocentrotus — представитель вторичноротых. Билатерии, по современным представлениям, вскоре после своего появления подразделились на две группы — первичноротых и вторичноротых. К первым относятся кольчатые, плоские и круглые черви, моллюски, членистоногие и много всякой мелочи, ко вторым — иглокожие, полухордовые и хордовые. В качестве «внешней» по отношению к билатериям группы был использован представитель кишечнополостных — актиния Nematostella (см.: Геном актинии оказался почти таким же сложным, как у человека, «Элементы», 11.07.2007).
Сравнивая геномы разных животных, ученые ранее установили, что у билатерий имеется свыше 30 уникальных микроРНК, которых нет у других живых организмов. Эти результаты подтвердились в ходе целенаправленного поиска микроРНК у платинереиса. У этого червя обнаружилось 34 микроРНК, общих для первично- и вторичноротых. Эти микроРНК уже имелись у последнего общего предка всех билатерий, но их не было у радиально-симметричных предков билатерий (кроме одной-единственной микроРНК, которая называется miR-100 и которая есть у актинии).
Таким образом, получается, что параллельно с резким усложнением организма и появлением новых тканей древние билатерии приобрели три десятка новых микроРНК. Главный вопрос состоит в том, была ли между этими новоприобретениями прямая связь. Если да, то следует ожидать, что в ходе онтогенеза у примитивных билатерий микроРНК распределяются неким закономерным образом по формирующимся тканям. Ранее это пытались изучить на «высших» животных, таких как членистоногие и хордовые, но картина получалась слишком сложная и запутанная.
Оказалось, что в ходе развития платинереиса микроРНК действительно очень четко распределены по дифференцирующимся тканям. В частности, древнейшая микроРНК miR-100 и три другие микроРНК обнаруживаются исключительно в двух небольших группах клеток по обе стороны от личиночной глотки (передней кишки). Клетки при ближайшем рассмотрении оказались нейросекреторными. Это интересно, потому что у позвоночных одна из этих молекул (miR-375) маркирует нейросекреторные клетки гипофиза и поджелудочной железы, формирование которых в онтогенезе связано с передней кишкой.
У актинии экспрессия miR-100 обнаружилась в отдельных клетках вдоль края личиночного рта (бластопора), примерно в той области, которая, по сравнительно-анатомическим данным, соответствует передней кишке или глотке билатерий (предполагается, что у предков билатерий был щелевидный рот, как у некоторых кишечнополостных, который потом склеился посередине, оставив два отверстия по краям, которые стали ртом и анусом).
У личинок червя Capitella и морского ежа эти микроРНК тоже локализуются вокруг глотки. По-видимому, такая их локализация первична для билатерий, но у продвинутых животных, таких как членистоногие и хордовые, эти микроРНК расширили сферу своей деятельности и стали работать не только в передней кишке и связанных с ней нейросекреторных клетках, но и в других частях зародыша.
Личинки кольчатых червей, иглокожих и многих других билатерий плавают при помощи ресничных шнуров — полосок эпителиальных клеток, покрытых согласованно бьющимися ресничками. Три микроРНК (miR-29, miR-34, miR-92) у обоих исследованных червей и морского ежа оказались приуроченными к этим ресничным шнурам. У позвоночных эти микроРНК экспрессируются в нейронах, выстилающих желудочки мозга, причем некоторые из этих нейронов несут реснички.
Этот факт должен привести в восторг сравнительных анатомов и эмбриологов, которые давно говорили о тесной связи между ресничными шнурами и нервными стволами и о том, что центральная нервная система хордовых формируется из спинной нервной трубки, которая образуется путем впячивания особой полоски эпителия, которая, в свою очередь, гомологична невротроху — ресничному шнуру, образующемуся у личинок билатерий на месте замкнувшегося щелевидного бластопора. Короче говоря, клетки с ресничками, выстилающие желудочки мозга позвоночных, исторически восходят к невротроху. Поэтому то, что в них экспрессируются те же микроРНК, что и в невротрохе, показывает, что сравнительные анатомы XIX–XX веков не даром ели свой хлеб. На приведенном рисунке невротрох личинки платинереиса (правда, без ресничек в данном случае) заметен в виде вертикальной полоски из красных точек на фигуре d.
|
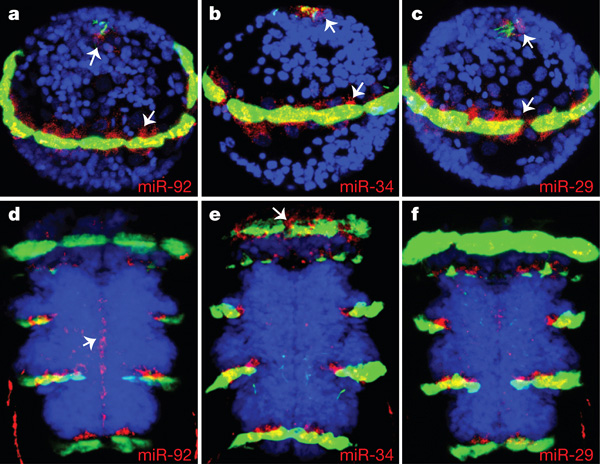
Два набора микроРНК оказались приуроченными к двум разным областям мозга. Один набор присутствует в нейросекреторной ткани дорсальной (спинной) части мозга. Те же самые микроРНК ранее были обнаружены в нейросекреторных клетках гипоталамуса у рыб. То, что гипоталамус находится в нижней («брюшной») части мозга, это правильно, потому что брюшная сторона хордовых гомологична спинной стороне других билатерий (В. В. Малахов. Происхождение хордовых животных, PDF, 236 Кб). Второй набор «мозговых» микроРНК у червей экспрессируется у оснований антенн — органов химического чувства. У мышей те же микроРНК приурочены к переднему мозгу, изначальной функцией которого у позвоночных была обработка обонятельной информации. Таким образом, здесь тоже наблюдается нечто весьма похожее на эволюционную преемственность.
Остальные микроРНК тоже оказались распределены закономерным образом по разным типам формирующихся тканей, причем были выявлены многочисленные параллели между их распределением у разных билатерий. Например, miR-124 приурочена к центральной нервной системе у насекомых и плоских червей, а у позвоночных — к нервным клеткам вообще. У платинереиса эта микроРНК экспрессируется только в центральной нервной системе — мозге и брюшной нервной цепочке. Другие микроРНК, у позвоночных экспрессирующиеся в некоторых органах чувств, у платинереиса тоже оказались приуроченными к чувствительным органам (глазам, антеннам, чувствительным придаткам параподий). МикроРНК miR-1 и miR-133 у кольчатых червей, как и у позвоночных, присутствуют только в развивающихся мышцах, и так далее.
Полученные результаты показывают, что одновременное появление у ранних билатерий новых тканей и новых микроРНК не было случайным совпадением. По-видимому, микроРНК изначально играли важную роль в дифференцировке тканей у билатерий. Их функции, однако, оказались более пластичными, чем у Hox-генов, и поэтому у высших билатерий «сфера влияния» многих микроРНК стала сильно отличаться от исходной.

")