|
Иммунитет помогает всем живым существам справляться с внедрением чужеродных агентов, в том числе и паразитов. Иммунная система — это сложный биохимический аппарат; у высших животных он нацелен на быстрое распознавание паразита и усиленное производство антител, которые помогают его обезвредить. Важнейшее звено иммунной машины позвоночных — выработка нужных, специфических, антител — является остроумным изобретением природы: из миллионов имеющихся лимфоцитов выбирается один или несколько таких, у которых поверхностные белки оказываются комплементарными антигену внедрившегося паразита. Образование комплекса антиген–антитело вызывает усиленное размножение именного этого типа лимфоцитов, что и обеспечивает быструю иммунную реакцию.
Источником замечательного разнообразия лимфоцитов служат неисчислимые комбинации нескольких коротких отрезков нуклеотидных последовательностей, из которых в созревающем лимфоците собираются гены антител. Таким образом, клетке не обязательно хранить отдельные гены для каждого антитела, можно хранить набор заготовок, а затем, по мере надобности, подбирать нужную комбинацию заготовок. Родители передают потомкам именно наборы заготовок. Поэтому приобретенный за жизнь иммунитет не может наследоваться, наследуется только возможность его приобрести. Так обстоят дела у позвоночных животных.
У беспозвоночных животных и растений иммунитет преимущественно врожденный (см. Иммунитет растений), а если и есть элементы приобретенного иммунитета, то развивающиеся в течение жизни организма защитные средства не передаются от родителей к потомкам — наследуется только способность их вырабатывать. А вот бактерии и археи, как выяснилось в исследованиях последних лет, способны передавать приобретенный иммунитет по наследству, демонстрируя тем самым один из редких случаев настоящего «ламарковского» наследования (см. Наследование приобретенных признаков).
В 2002 году началось планомерное изучение особых участков (локусов) бактериального генома, которые представляют собой расположенные группами короткие палиндромные повторы (CRISPR, clustered regularly interspaced short palindromic repeats). Эти локусы найдены у 90% архей и 40% бактерий.
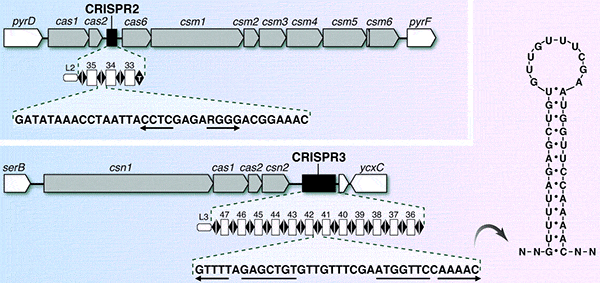
Локусы CRISPR состоят из нескольких несоприкасающихся палиндромных повторов, между которыми имеются промежутки — спейсеры. Спейсер представляет собой короткий участок вирусной или плазмидной ДНК. Размер CRISPR-повтора исчисляется 23–47 нуклеотидными парами, а спейсеров — от 21 до 72 нуклеотидных пар. Число групп «повтор/спейсер» может достигать 375, но обычно меньше 50. В бактериальном геноме может быть не один, а несколько локусов CRISPR.
В непосредственной близости от CRISPR расположены гены специальных белков, называемых Cas (CRISPR associated). Обычно Cas — это нуклеазы, полимеразы, нуклеотид-связывающие белки; всего эта группа объединяет около 40 семейств белков.
Повторы CRISPR-последовательностей очень консервативны в пределах каждого вида микробов, но сильно различаются от вида к виду.
|
В 2007 году было экспериментально показано, что бактерия Streptococcus thermophilus в ходе борьбы с вирусами-бактериофагами изменяет последовательность CRISPR, достраивая к концу старой последовательности еще одну или несколько единиц «повтор/спейсер». Добавленные спейсеры были аналогичны участкам вирусного генома. При этом бактерии приобретали устойчивость к данному вирусу. Если изменить последовательность нуклеотидов добавленного спейсера или вообще вырезать спейсер, то приобретенная устойчивость к фагу теряется.
В последующие два года ученые доказали, что локусы CRISPR эволюционируют в ответ на вирусную активность, так что состав и порядок расположения спейсеров указывает на историю различных вирусных атак. Итак, взаимодействие с вирусами и чужеродными плазмидами приводит к появлению иммунитета, который сохраняется в ряде поколений данного штамма. Ученые предположили, каким может быть механизм приобретения и наследования иммунитета.
Во-первых, в формировании иммунитета участвуют Сas-белки. Инактивация cas-генов ведет к снижению или потере способности интегрировать в качестве спейсеров вирусную или плазмидную ДНК. Если в клетку проникает чужеродная ДНК, то Сas-белки перво-наперво распознают ее. Потом они же вырезают участок чужеродной ДНК и достраивают в локус CRISPR новую рабочую единицу «повтор/спейсер». Обновленная бактериальная ДНК в дальнейшем нормально дуплицируется и передается по наследству потомкам данной бактерии. При транскрипции CRISPR формируется цепочка РНК (CRISPR-РНК, или crРНК), которая затем нарезается на короткие кусочки, состоящие из двух половинок палиндромного повтора и заключенного между ними спейсера. Эту нарезку также осуществляют Сas-белки.
В результате получается внушительный набор коротких crРНК с различными вирусными спейсерами. Среди них и тот, который был счастливо приобретен при недавнем инфицировании. crРНК объединяются с некоторыми Сas-белками. Если в такую клетку снова попадает этот вирус, то РНК, несущая соответствующий спейсер, узнает комплементарный участок вирусной ДНК, а Сas-белки обеспечивают инактивацию и избавление от паразитической ДНК. Само собой разумеется, что узнавание чужеродной ДНК при помощи crРНК осуществляется гораздо эффективнее и быстрее, чем исходное распознавание, с которого началось формирование иммунитета.
Можно себе представить, что при частых атаках определенного вируса соответствующая рабочая единица «повтор/спейсер» будет поддерживаться в рабочем состоянии в ряде поколений (мутации, повреждающие ее, будут отсеиваться отбором). Если же паразит оказался редким или экзотическим, то соответствующая единица «повтор/спейсер» быстро накопит мутации, перестанет работать или исчезнет вовсе. Так что CRISPR-иммунитет — это самонастраивающаяся система. Взаимодействие между бактериями и вирусами заставляет их геномы эволюционировать чрезвычайно быстро.
Совсем недавно американские ученые из Северо-Западного университета (Эванстон, Иллинойс) открыли, как бактериям удается избежать аутоиммунной реакции при функционировании crРНК. Гипотетически crРНК должны бы распознать участок «родного» локуса CRISPR в бактериальной хромосоме и деактивировать его. Но ведь этого не происходит. Оказалось, что в распознавании чужеродных ДНК участвуют не только спейсеры, но и ограничивающие спейсер фрагменты палиндромных повторов. Если при спаривании crРНК с ДНК комплементарным оказывается не только спейсер, но и окружающие его нуклеотиды, то crРНК узнает «родную» ДНК, и иммунная атака не происходит. Если же спаривается только спейсер crРНК, то это означает чужеродную цепочку, и тогда белки Cas избавляются от паразитической ДНК.

")