Биологи из Торонтского университета (Канада) провели на дрожжах опыты по искусственному видообразованию. Ранее в подобных экспериментах пытались (и порой успешно) получить поведенческую презиготическую изоляцию, то есть нежелание представителей разошедшихся линий скрещиваться друг с другом. На этот раз удалось получить постзиготическую изоляцию, то есть снижение жизнеспособности гибридов. Дрожжи, выращивавшиеся в течение 500 поколений в контрастных неблагоприятных условиях высокой солености или низкого содержания глюкозы, оказались постзиготически изолированы как друг от друга, так и от предковой линии.
|
Хотя в природе есть сколько угодно «хороших видов», способных скрещиваться друг с другом и давать плодовитое гибридное потомство, главным критерием вида для организмов с половым размножением все-таки считается репродуктивная изоляция. Именно на ней и сосредотачиваются усилия биологов, ставящих эксперименты по искусственному видообразованию.
В большинстве подобных экспериментов разные линии подопытных организмов подвергают разнонаправленному отбору (например, помещают в контрастные неблагоприятные условия и выращивают в течение нескольких десятков поколений), а затем смотрят, не отбило ли у них охоту к перекрестному скрещиванию. Надо сказать, что иногда охоту действительно отбивает, животные начинают предпочитать в качестве брачных партнеров «своих», то есть тех, чьи предки подвергались такому же отбору, приспосабливались к таким же условиям (см.: Опыты по искусственному видообразованию). В этих экспериментах изучается зарождение презиготической изоляции (изоляции до оплодотворения, до образования зиготы).
Но изоляция может быть и постзиготической. Это значит, что особи спариваются, оплодотворение происходит, но гибридное потомство либо погибает, либо оказывается стерильным, либо имеет резко пониженную жизнеспособность. В такой ситуации скрещиваться с чужаками становится крайне невыгодно, и это, в свою очередь, может стать предпосылкой для постепенной выработки презиготической изоляции (отбор будет благоприятствовать особям, спаривающимся только со «своими»).
Экспериментов, демонстрирующих развитие постзиготической изоляции, проведено очень мало. Авторы сообщают, что им известна только одна такая работа, выполненная на дрозофиле в 1980 году (Oliveira, Cordeiro. «Adaptation of Drosophila willistoni experimental populations to extreme pH medium II. Development of incipient reproductive isolation» // Heredity. 1980. V. 44. P. 123–130). Странно, что не упомянуты работы советского энтомолога Георгия Христофоровича Шапошникова, который получил сходные результаты в экспериментах с тлями (Шапошников Г.Х. «Морфологическая дивергенция и конвергенция в эксперименте с тлями (Homoptera, Aphidenea)» // Энтомол. обозр. 1965. Т. 44. № 1. С. 3–25), но так уж повелось: советские (а теперь российские) научные журналы на Западе читают редко.
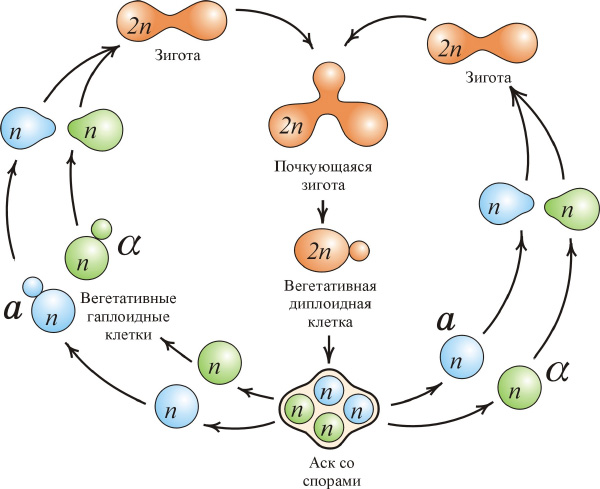
Исследователи выбрали в качестве объекта дрожжи Saccharomyces cerevisae. Подобно тлям, дрожжи размножаются как половым путем, так и бесполым (см. рис.), с той разницей, что бесполое размножение у тлей осуществляется путем партеногенеза (развитие из неоплодотворенных яиц), а у дрожжей — путем почкования. Из одной родительской диплоидной клетки было произведено 12 подопытных популяций. Изначально все они были генетически идентичны, но потом в результате мутаций и отбора могли накапливать различия. Популяции выращивали в течение 500 поколений в контрастных неблагоприятных условиях: 6 популяций при повышенной солености, 6 — в «минимальной» среде с низким содержанием глюкозы. Контрольную (предковую) линию продолжали растить в нормальной среде.
После этого ученые проверили, произошла ли адаптация, то есть стали ли подопытные популяции лучше приспособлены к своим средам по сравнению с предками. Приспособленность оценивалась по скорости бесполого размножения (почкования) клеток в условиях конкуренции с предковой линией. Например, чтобы измерить приспособленность «соленой» популяции (S), в соленую среду помещали равное количество «соленых» и предковых (P) клеток, а через сутки смотрели, какие клетки большее число раз успели поделиться.
Оказалось, что адаптация действительно произошла во всех 12 линиях, хотя величина приспособленности варьировала. К соленой среде дрожжи в целом приспособились несколько лучше, чем к голодной.
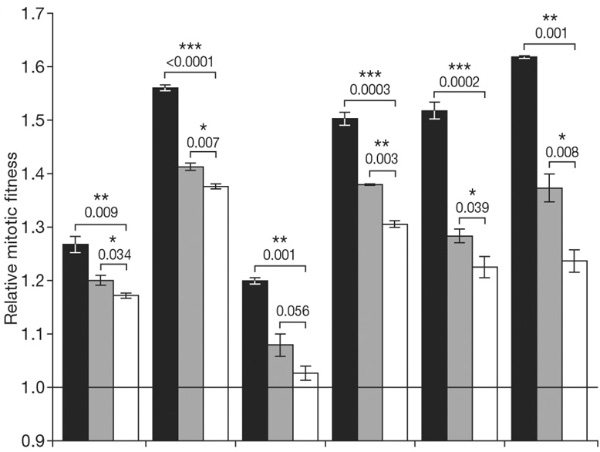
Теперь можно было приступить к самой главной части эксперимента — к выявлению постзиготической изоляции, если таковая возникла. Исследователи получили гибридное потомство, скрестив каждую из линий S с одной из «голодных» линий (M). Всего, таким образом, получилось 6 линий «солено-голодных» гибридов S/M. Кроме того, были получены гибриды каждой из 12 линий с предками (гибриды S/P и M/P). Оказалось, что чистые линии S и M приспособлены к своим средам значительно лучше, чем любые гибриды. Стало быть, постзиготическая изоляция налицо.
|
Пониженная жизнеспособность гибридов S/M проявилась также и в резком (примерно в два раза) снижении «мейотической эффективности», то есть способности к образованию спор в подходящих для этого условиях.
Наличие разнообразных гибридных линий дало возможность понять некоторые детали тех механизмов, которые обусловили пониженную жизнеспособность гибридов.
Если приспособленность популяций к своим средам была обусловлена только лишь накоплением полезных мутаций, проявляющихся независимо друг от друга, то приспособленность гибридов разного типа должна быть примерно одинаковой. Например, в соленой среде гибриды S/M и S/P должны демонстрировать одинаковую приспособленность (или неприспособленность). Это, однако, оказалось не так. Гибриды S/P в соленой среде чувствовали себя заметно лучше, чем S/M (хотя и намного хуже, чем чистая линия S). То же самое, хотя и в меньшей степени, справедливо и для «голодной» среды: здесь гибриды M/P оказались более приспособленными, чем S/M, в 5 случаях из 6, но только в трех случаях различие оказалось статистически значимым.
Этот результат можно объяснить двояко. Либо адаптации к одной из двух неблагоприятных сред сами по себе снижают приспособленность к альтернативной среде (detrimental pleiotropic effects of adaptation on fitness in the alternative environment), либо имеет место так называемый антагонистический эпистаз (antagonistic epistasis) между компонентами геномов S и M, то есть гены, ответственные за адаптацию к разным средам, каким-то образом мешают друг другу работать. В первом случае дрожжи из линии S должны расти в голодной среде хуже, чем P, а в соленой среде P должны расти лучше, чем M. Это не подтвердилось, следовательно главную роль в снижении приспособленности S/M по сравнению с S/P и M/P играет антагонистический эпистаз.
Исследование наглядно показало, что разнонаправленный отбор может приводить к довольно быстрому формированию неполной постзиготической репродуктивной изоляции.
Чрезвычайно интересно было бы узнать, одинаковые или разные приспособления возникли в линиях дрожжей, параллельно приспособившихся к одинаковой среде (например, у шести «соленых» популяций). К сожалению, в статье нет четкого ответа на этот вопрос. Правда, там отмечено, что гибриды между разными линиями S (как и между разными линиями M) достоверно не отличались по приспособленности от чистых линий S и M. Это свидетельствует в пользу того, что адаптации возникли во многом сходные, по крайней мере совместимые и взаимозаменимые. С другой стороны, достигнутый разными линиями уровень приспособленности заметно различался.

")